§ 22-1. Цемнавая фаза фотасінтэзу. Значэнне фотасінтэзу. Хемасінтэз
Цемнавая фаза *(на прыкладзе С3-фотасінтэзу). Як вы ведаеце, рэакцыі цемнавой фазы ажыццяўляюцца ў строме хларапласту незалежна ад наяўнасці святла. Аднак для іх працякання неабходны такія прадукты светлавой фазы, як АТФ адноўлены НАДФ.*
З навакольнага асяроддзя ў хларапласты паступае вуглякіслы газ. Пад час цёмнай фазы фотасінтэзу адбыва*звязванне (фіксацыя) і* аднаўленне да арганічных рэчываў. *У фотааўтатрофаў выяўлена некалькі розных спосабаў фіксацыі СО2 і яго наступнага выкарыстання для сінтэзу арганічных злучэнняў. Пры гэтым асноўным, найбольш пашыраным, механізмам з'яўляецца так званы С3-шлях фотасінтэзу. Яго можна падзяліць на некалькі этапаў.
1. Звязванне вуглякіслага газу. У строме хларапласту малекулу СО2 далучае асаблівы акцэптар — рыбулоза-1,5-дыфасфат (РДФ). Гэта вытворнае пяцівугляроднага монацукрыду рыбулозы. Рэакцыя працякае з выкарыстаннем вады і каталізуецца ферментам РДФ-карбаксілазай, на долю якой можа прыпадаць больш за палову ўсіх бялкоў хларапластаў. Лічыцца, што РДФ-карбаксілаза — самы пашыраны бялок на Зямлі. У выніку далучэння вуглякіслага газу да РДФ утвараецца няўстойлівае шасцівугляроднае злучэнне, якое распадаецца на дзве малекулы фосфагліцэрынавай кіслаты (ФГК):

Малекула ФГК — першаснага прадукта фіксацыі СО2 — змяшчае 3 атамы вугляроду. Таму дадзены механізм злучэння вуглякіслага газу і названы С3-шляхам фотасінтэзу.
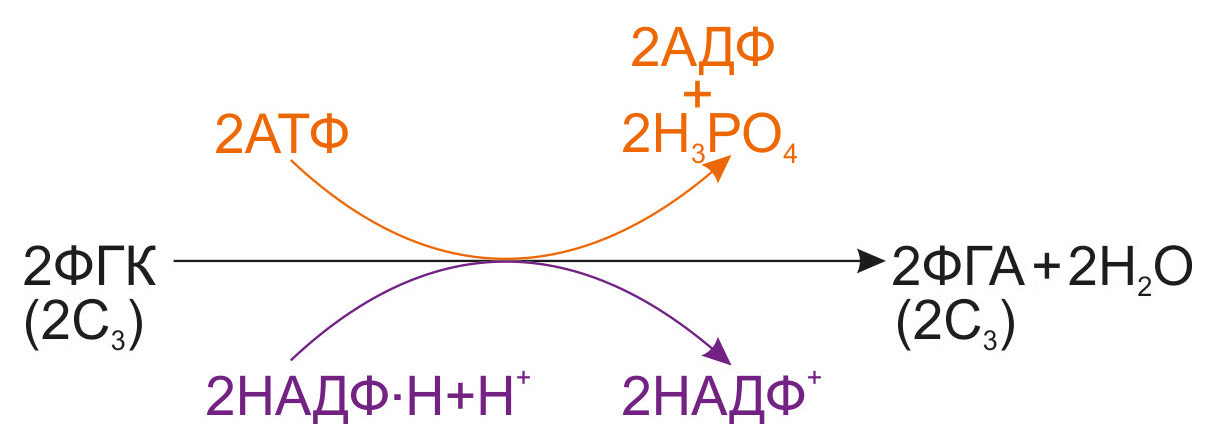
2. Аднаўленне ФГК. Далей фосфагліцэрынавая кіслата падягае ферментацыйнаму аднаўленню да фосфагліцэрынавага альдэгіду (ФГА). Працэс працякае з выкарыстаннем прадуктаў светлавой фазы фотасінтэзу — АТФ и НАДФ∙Н+Н+:
3. Ператварэнне ФГА ў іншыя прадукты фотасінтэзу і РДФ. Некаторая частка малекул ФГА ідзе на сінтэз глюкозы і іншых монацукрыдаў, спіртоў, карбонавых кіслот, амінакіслот (утвараюцца шляхам амінавання карбонавых кіслот) і г. д. Далей з іх могуць сінтэзавацца больш складаныя злучэнні — аліга- і поліцукрыды (крухмал, цэлюлоза), ліпіды, бялкі і інш.
Аднак большая частка малекул ФГА ператвараецца ў рыбулоза-1,5-дыфасфат, здольны зноў звязваць СО2. Рэакцыі рэгенерацыі РДФ працякаюць з затратамі энергіі АТФ. Такім чынам, С3-шлях фіксацыі вуглякіслага газу ўяўляе сабой цыклічны працэс. У гонар амерыканскага біяхіміка М. Кальвіна, які даследаваў гэты спосаб асіміляцыі СО2 (Нобелеўская прэмія за 1961 г.), ён названы цыклам Кальвіна.*
*За адзін «абарот» цыкла Кальвіна фіксуецца адна малекула вуглякіслага газу. Таму для сінтэзу шасцівугляроднай малекулы глюкозы (С6Н12О6) патрабуецца шэсць «абаротаў» цыкла. Пяць з іх неабходны для рэгенерацыі РДФ, а адзін ідзе ўласна на ўтварэнне глюкозы (мал. 22-1.1).*

*Пры С3-фотасінтэзе для ўтварэння адной малекулы глюкозы з СО2 неабходна выкарыстоўваць, а потым зноў рэгенераваць 6 малекул РДФ, акісліць 12 малекул НАДФ∙Н+Н+ (ён служыць крыніцай атамаў вадароду) і расшчапіць 18 малекул АТФ (яна з'яўляецца пастаўшчыком энергіі). Агульнае ўраўненне цемнавой фазы фотасінтэзу можна запісаць наступным чынам (для спрашчэння не паказаны малекулы дыгідрамонааксіду, патрэбныя для гідролізу АТФ):
6СО2 + 12НАДФ∙Н+Н+ + 18АТФ → С6Н12О6 + 6Н2О + 12НАДФ+ + 18АДФ + 18Н3РО4.*
Такім чынам, АТФ * і адноўлены НАДФ, атрыманыя ў ходзе светлавой фазы, выкарыстоўваюцца ў цемнавой фазе для ўтварэння глюкозы і іншых прадуктаў фотасінтэзу. Пры гэтым энергія макраэргетычных сувязей АТФ пераўтворыцца ў энергію хімічных сувязей арганічных рэчываў.
*Як ужо адзначалася, рэакцыі цемнавой фазы працякаюць амаль адначасова са светлавой. Эксперыменты паказалі, што на святле ў хларапластах адбываецца звязванне вуглякіслага газу і сінтэз глюкозы. Пасля выключэння святла ўтварэнне глюкозы пэўны час працягваецца, а потым спыняецца. Але, калі ў асяроддзе з хларапластамі дабавіць АТФ і НАДФ∙Н+Н+, сінтэз глюкозы адновіцца. Ён будзе працякаць у цемнаце датуль, пакуль не вычарпаюцца запасы АТФ і адноўленага НАДФ.*
*Такім чынам, для сінтэзу малекулы глюкозы трэба 12 малекул НАДФ∙Н+Н+. Каб аднавіць 12 малекул НАДФ+ у светлавой фазе, неабходна далучыць да іх 24 атамы вадароду, г. зн. 24 электроны (е–) і 24 пратоны (Н+). А для іх утварэння патрабуецца ажыццявіць фатоліз 12 малекул вады:
12Н2О → 6О2 + 24е– + 24Н+.*
Калі аб'яднаць працэсы, якія праходзяць у светлавой і цемнавой фазах, выключыўшы прамежкавыя прадукты * і скараціўшы малекулы вады (мал. 22-1.2)*, можна атрымаць сумарнае ўраўненне фотасінтэзу:


*Іншыя шляхі фіксацыі СО2 пры фотасінтэзе. У некаторых раслін першасным прадуктам фіксацыі вуглякіслага газу з'яўляецца не ФГК, як пры С3-шляху фотасінтэзу, а чатырохвугляроднае злучэнне — шчаўевавоцатная кіслата (ШВК). Такі механізм звязвання СО2 вядомы як С4-шлях фотасінтэзу, ці шлях Хэтча–Слэка (у гонар аўстралійскіх вучоных М. Д. Хэтча і Ч. Р. Слэка, якія дэталёва даследавалі гэты спосаб асіміляцыі вуглякіслага газу).
Для раслін, якія выкарыстоўваюць С4-шлях фотасінтэзу (напрыклад, кукурузы, проса, сорга, цукровага трыснягу), характэрна асаблівая будова лістоў. Іх праводзячыя пучкі акружаны двума слаямі фотасінтэзуючых клетак. Унутраны слой утварае так званую абкладку праводзячага пучка. Вонкавы слой прадстаўлены клеткамі мезафілу (асноўнай хларафіланоснай тканкі лістоў раслін, у большасці выпадкаў дыферэнцыяванай на слупкаватую і губчатую парэнхіму), у гіялаплазме якіх і адбываецца фіксацыя СО2 па С4-шляху.
Акцэптарам вуглякіслага газу пры С4-фотасінтэзе служыць трохвугляроднае злучэнне — фосфаэнолпіравінаградная кіслата (ФЕП):

Вызначана, што ФЕП-карбаксілаза, якая каталізуе гэту рэакцыю, больш актыўна звязвае СО2, чым РДФ-карбаксілаза, што выконвае такую ж функцыю ў цыкле Кальвіна.
Шчаўевавоцатная кіслата, утвораная ў клетках мезафілу, ператвараецца ў яблычную (ці аспарагінавую) кіслату, якая паступае ў клеткі абкладкі праводзячага пучка (мал. 22-1.3). Тут шляхам адшчаплення малекулы СО2 яна ператвараецца ў ПВК. У клетках абкладкі ажыццяўляецца стандартны С3-шлях злучэння вуглякіслага газу, г. зн. выдзелены СО2 трапяе ў цыкл Кальвіна. ПВК вяртаецца ў клеткі мезафілу, дзе ператвараецца ў ФЕП — зыходны акцэптар вуглякіслага газу.*

Вядома, што ў С3-раслін, якія фіксуюць СО2 толькі з дапамогай цыкла Кальвіна, фотасінтэз працякае хутка толькі пры даволі высокай канцэнтрацыі вуглякіслага газу ў паветры. С4-шлях нават ва ўмовах ўтрымання СО2 дазваляе незапашваць у клетках абкладкі вуглякіслы газ у колькасці, дастатковай для эфектыўнага працякання цыкла Кальвіна. Такім чынам, у С4-раслін фотасінтэз ажыццяўляецца інтэнсіўна і пры нізкіх канцэнтрацыях СО2 у навакольным асяроддзі. Пры гэтым адпадае неабходнасць ўвесь час трымаць вусцейкі адкрытымі (для паступлення большай колькасці СО2 да фотасінтэзуючых клетак), што зніжае страты вады у ходзе транспірацыі. Усё гэта дазваляе С4-раслінам асвойваць засушлівыя, гарачыя месцапражывання.
У падобных умовах могуць існаваць і расліны, якія выкарыстоўваюць так званы САМ-фотасінтэз (скарачэнне ад англ. crassulacean acid metabolism — кіслотны метабалізм таўсцянкавых). Гэты шлях характэрны пераважна для сукулентаў — таўсцянак, кактусаў, каланхоэ і інш. У сувязі са строгай эканоміяй вады вусцейкі САМ-раслін у дзённы час закрыты. Адкрываюцца яны толькі ноччу, у самы халаднаваты перыяд сутак. Такім чынам, вуглякіслы газ можа паступаць у лісты толькі ноччу. У гіялаплазме хларафіланосных клетак ён фіксуецца па С4-шляху і назапашліваецца ў вакуолях у выглядзе яблычнай кіслаты (гл. мал. 22-1.3). Удзень, калі вусцейкі закрыты, яблычная кіслата пераходзіць у гіялаплазму і вызваляе назапашаны С2. Ён паступае ў строму хларапластаў і ўключаецца ў цыкл Кальвіна.
Такім чынам, механізм САМ-фотасінтэзу падобны да С4-фотасінтэзу. Адрозненне заключаецца ў тым, што ў С4-раслін звязванне вуглякіслага газу па С4- і С3-шляху адбываецца ў розных тыпах клетак, а ў САМ-раслін — у межах адной і той жа клеткі, але ў розны час сутак.*