§ 33. Дыгібрыднае скрыжаванне. Трэці закон Мендэля
Цыталагічныя асновы закона незалежнага наследавання прымет. У разгледжаным эксперыменце Г. Мендэль вывучаў наследаванне дзвюх пар альтэрнатыўных прымет. Відавочна, што колер насення і форма яго паверхні вызначаюцца дзвюма рознымі парамі генаў. Абазначым алель жоўтай афарбоўкі А, зялёнай — а, гладкай формы — в, маршчыністай — b.
Гены, якія кантралююць розныя пары прымет (г. зн. неальтэрнатыўныя прыметы), называюцца неалельнымі. Яны размяшчаюцца ў розных парах храмасом ці ў розных участках (локусах) гамалагічных храмасом. У дадзеным выпадку гены, якія вызначаюць афарбоўку (А і а), неалельныя ў адносінах да генаў, што абумоўліваюць форму паверхні насення (В і b). Уявім, што гэтыя пары алеляў знаходзяцца ў негамалагічных храмасомах, г. зн. у розных парах храмасом.
Бацькоўскія расліны маюць генатыпы AABB і aabb. Арганізмы, гомазіготныя па дзвюх парах генаў, называюцца дыгомазіготамі. У гаметы трапляе па адным гене з кожнай пары. Значыць, у дыгомазіготных бацькоў фарміруецца толькі адзін тып гамет: у аднаго — AB, у другога — ab. У выніку апладнення развіваецца першае пакаленне гібрыдаў. Усе яны маюць жоўтае гладкае насенне, што абумоўлена генатыпам AaBb. Асобіны, гетэразіготныя па дзвюх парах генаў, называюцца дыгетэразіготамі. Колькі тыпаў гамет утвараюць дыгетэразіготныя асобіны?
Вам вядома, што ў анафазе I меёзу гамалагічныя храмасомы расцягваюцца да розных полюсаў клеткі. Пры гэтым разыходжанне кожнай пары храмасом адбываецца незалежна ад іншых пар. Негамалагічныя храмасомы разыходзяцца да полюсаў выпадкова, утвараючы розныя камбінацыі (мал. 33.2). Такім чынам, ген А можа апынуцца ў адной палавой клетцы з генам В ці з генам b. Менавіта гэтак жа ген а можа трапіць у адну гамету з генам В або з генам b. У выніку гэтага дыгетэразіготныя асобіны ўтвараюць чатыры тыпы гамет: AB, Ab, aB, ab у роўных суадносінах — па 25 %.

Выпадковае зліццё палавых клетак пры апладненні вядзе да ўтварэння розных тыпаў зігот, а значыць, і нашчадкаў. Выкарыстоўваючы фенатыпічныя радыкалы, расшчапленне па фенатыпе ў другім гібрыдным пакаленні (пры ўмове, што алелі кожнай пары ўзаемадзейнічаюць па тыпе поўнага дамінавання) можна запісаць у выглядзе:
9A—B— : 3A—bb : 3aaB— : 1aabb.
*Тое, што разгледжанае дыгібрыднае скрыжаванне ўяўляе сабой два монагібрыдныя, якія ідуць незалежна, можна прадэманстраваць наступным чынам:
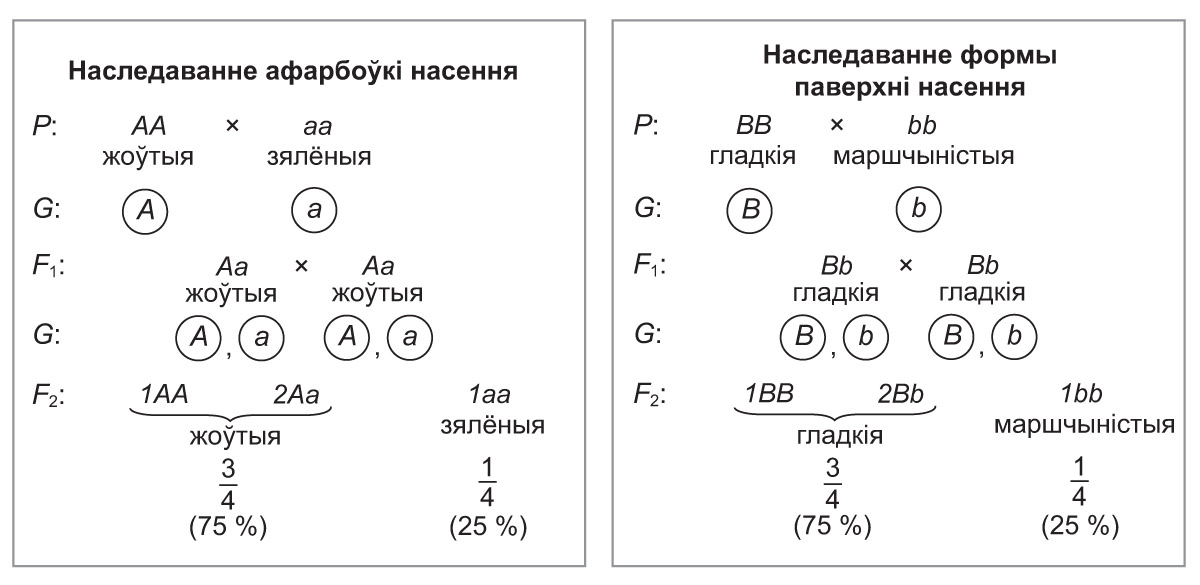
На аснове монагібрыдных скрыжаванняў вызначым вынікі дыгібрыднага. У другім пакаленні долі асобін з рознымі прыметамі насення складаюць:
● жоўтае гладкае =  жоўтае × гладкае =
жоўтае × гладкае =  (або 75 % × 75 % : 100 % = 56,25 %);
(або 75 % × 75 % : 100 % = 56,25 %);
● жоўтае маршчыністае = жоўтае ×  маршчыністае =
маршчыністае =  (або 75 % × 25 % : 100 % = 18,75 %);
(або 75 % × 25 % : 100 % = 18,75 %);
● зялёнае гладкае = зялёнае × гладкае = (або 25 % × 75 % : 100 % = 18,75 %);
● зялёнае маршчыністае = зялёнае × маршчыністае =  (або 25 % × 25 % : 100 % = 6,25 %).*
(або 25 % × 25 % : 100 % = 6,25 %).*
Такім чынам, у аснове незалежнага наследавання ляжыць:
1) выпадковае разыходжанне негамалагічных храмасом у анафазе I меёзу, якое прыводзіць да ўтварэння гамет з рознымі камбінацыямі генаў;
2) выпадковае зліццё гамет пры апладненні, што абумоўлівае фарміраванне розных тыпаў зігот.
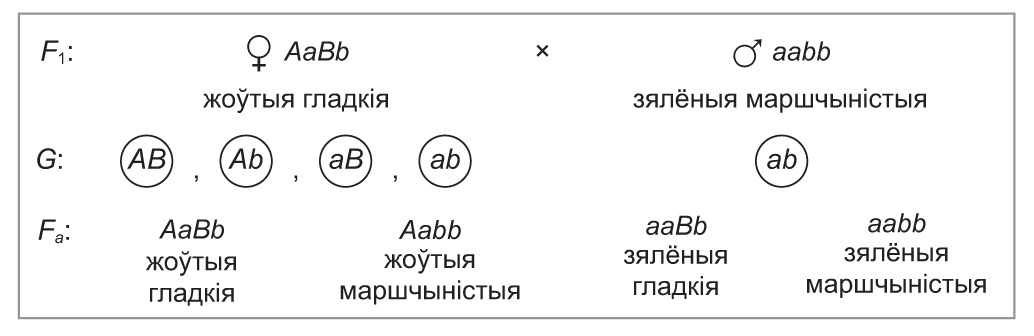
Даказаць, што ў дыгетэразіготнай асобіны ўтвараюцца палавыя клеткі чатырох тыпаў, прычым у роўных суадносінах, можна з дапамогай аналізуючага скрыжавання. Для гэтага дыгетэразіготны гарох, які мае жоўтае гладкае насенне, скрыжуем з рэцэсіўнай дыгомазіготай. У патомстве будзе назірацца чатыры фенатыпічныя і генатыпічныя класы ў суадносінах 1 : 1 : 1 : 1, г. зн. па 25 %. Гэта сведчыць пра роўнаверагоднае фарміраванне чатырох тыпаў гамет у таго з бацькоў, якога аналізуюць.