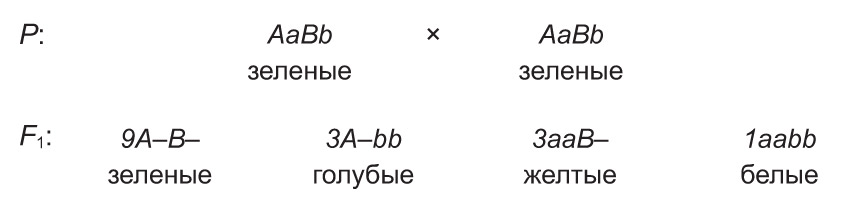*§ 33-1. Взаимодействие неаллельных генов
Признаки, наследование которых мы рассматривали ранее, контролируются генами одной аллельной пары. Однако многие признаки живых организмов определяются взаимодействием двух или более пар генов, т. е. неаллельными генами. Основными типами взаимодействия неаллельных генов являются комплементарность, эпистаз и полимерия.
Комплементарность выражается в том, что доминантные гены разных аллелей совместно обусловливают появление нового признака, не определявшегося ни одним из этих генов по отдельности. Например, у льна доминантный ген одной аллели (А) определяет розовую окраску цветков, а доминантный ген другой аллели (В) — белую. Если в генотипе растения присутствуют оба доминантных аллеля, его цветки имеют голубую окраску. Таким образом, неаллельные гены А и В действуют комплементарно (взаимно дополняя друг друга), что приводит к формированию нового признака. Суть такого взаимодействия можно отразить в виде схемы.

Как видно из схемы, синтез голубого пигмента из неокрашенного предшественника (пропигмента) осуществляется в два этапа, каждый из которых катализируется особым ферментом. Присутствие этих ферментов в клетках лепестков льна определяется наличием в генотипе соответствующих генов — А и В. Так, ген А определяет наличие фермента, который превращает пропигмент в промежуточный продукт — пигмент розового цвета. Ген В обусловливает присутствие второго фермента, синтезирующего голубой пигмент из розового. Поэтому у растений, имеющих оба доминантных аллеля, в лепестках образуется конечный продукт — пигмент голубого цвета. Если отсутствует ген В (а значит, и соответствующий фермент), в клетках протекает лишь первая стадия — образование розового пигмента. В случае отсутствия гена А (или обоих доминантных аллелей) не осуществляется даже первый этап синтеза пигмента, и лепестки остаются белыми.
Рассмотрим скрещивание дигомозиготных растений льна, имеющих розовые (AAbb) и белые (aaBB) цветки (рис. 33-1.1). В первом поколении наблюдается единообразие гибридов. Все они имеют голубые цветки, что обусловлено наличием в генотипе как гена А, так и гена В. Во втором гибридном поколении происходит расщепление в соотношении 9 голубых : 3 розовых : 4 белых.

Комплементарное взаимодействие генов лежит в основе проявления ряда признаков у различных организмов. Так, по типу комплементарности взаимодействуют гены, контролирующие окраску коконов у тутового шелкопряда и цветков у душистого горошка, форму гребня у кур, цвет оперения у попугаев и т. д. В зависимости от особенностей фенотипического проявления генов в потомстве от скрещивания дигетерозигот могут наблюдаться различные варианты расщепления — 9 : 7, 9 : 3 : 4, 9 : 6 : 1, 9 : 3 : 3 : 1.
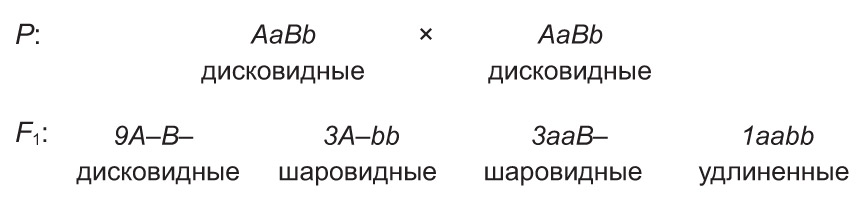
Например, у тыквы доминантные аллели А и В совместно приводят к появлению плодов дисковидной формы, а по отдельности — шаровидной. В то же время рецессивные аллели этих генов обусловливают удлиненную форму плодов. При скрещивании дигетерозиготных растений, имеющих дисковидные плоды, в потомстве происходит расщепление 9 : 6 : 1.
Сходным образом наследуется цвет оперения волнистых попугаев. Доминантный ген одной аллели (А) обусловливает у них голубую окраску перьев, а доминантный ген другой аллели (В) — желтую. При наличии в генотипе обоих доминантных аллелей (А и В) проявляется новый признак — зеленое оперение. Вместе с тем рецессивные аллели этих генов (a и b) определяют белую окраску перьев. Поэтому скрещивание зеленых дигетерозиготных особей приводит к расщеплению по цвету оперения среди потомков в соотношении 9 : 3 : 3 : 1.