*§ 33-1. Взаимодействие неаллельных генов
| Сайт: | Профильное обучение |
| Курс: | Биология. 11 класс |
| Книга: | *§ 33-1. Взаимодействие неаллельных генов |
| Напечатано:: | Гость |
| Дата: | Monday, 15 June 2026, 14:07 |
Признаки, наследование которых мы рассматривали ранее, контролируются генами одной аллельной пары. Однако многие признаки живых организмов определяются взаимодействием двух или более пар генов, т. е. неаллельными генами. Основными типами взаимодействия неаллельных генов являются комплементарность, эпистаз и полимерия.
Комплементарность выражается в том, что доминантные гены разных аллелей совместно обусловливают появление нового признака, не определявшегося ни одним из этих генов по отдельности. Например, у льна доминантный ген одной аллели (А) определяет розовую окраску цветков, а доминантный ген другой аллели (В) — белую. Если в генотипе растения присутствуют оба доминантных аллеля, его цветки имеют голубую окраску. Таким образом, неаллельные гены А и В действуют комплементарно (взаимно дополняя друг друга), что приводит к формированию нового признака. Суть такого взаимодействия можно отразить в виде схемы.

Как видно из схемы, синтез голубого пигмента из неокрашенного предшественника (пропигмента) осуществляется в два этапа, каждый из которых катализируется особым ферментом. Присутствие этих ферментов в клетках лепестков льна определяется наличием в генотипе соответствующих генов — А и В. Так, ген А определяет наличие фермента, который превращает пропигмент в промежуточный продукт — пигмент розового цвета. Ген В обусловливает присутствие второго фермента, синтезирующего голубой пигмент из розового. Поэтому у растений, имеющих оба доминантных аллеля, в лепестках образуется конечный продукт — пигмент голубого цвета. Если отсутствует ген В (а значит, и соответствующий фермент), в клетках протекает лишь первая стадия — образование розового пигмента. В случае отсутствия гена А (или обоих доминантных аллелей) не осуществляется даже первый этап синтеза пигмента, и лепестки остаются белыми.
Рассмотрим скрещивание дигомозиготных растений льна, имеющих розовые (AAbb) и белые (aaBB) цветки (рис. 33-1.1). В первом поколении наблюдается единообразие гибридов. Все они имеют голубые цветки, что обусловлено наличием в генотипе как гена А, так и гена В. Во втором гибридном поколении происходит расщепление в соотношении 9 голубых : 3 розовых : 4 белых.

Комплементарное взаимодействие генов лежит в основе проявления ряда признаков у различных организмов. Так, по типу комплементарности взаимодействуют гены, контролирующие окраску коконов у тутового шелкопряда и цветков у душистого горошка, форму гребня у кур, цвет оперения у попугаев и т. д. В зависимости от особенностей фенотипического проявления генов в потомстве от скрещивания дигетерозигот могут наблюдаться различные варианты расщепления — 9 : 7, 9 : 3 : 4, 9 : 6 : 1, 9 : 3 : 3 : 1.
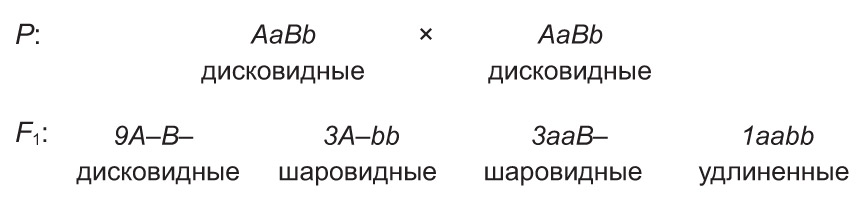
Например, у тыквы доминантные аллели А и В совместно приводят к появлению плодов дисковидной формы, а по отдельности — шаровидной. В то же время рецессивные аллели этих генов обусловливают удлиненную форму плодов. При скрещивании дигетерозиготных растений, имеющих дисковидные плоды, в потомстве происходит расщепление 9 : 6 : 1.
Сходным образом наследуется цвет оперения волнистых попугаев. Доминантный ген одной аллели (А) обусловливает у них голубую окраску перьев, а доминантный ген другой аллели (В) — желтую. При наличии в генотипе обоих доминантных аллелей (А и В) проявляется новый признак — зеленое оперение. Вместе с тем рецессивные аллели этих генов (a и b) определяют белую окраску перьев. Поэтому скрещивание зеленых дигетерозиготных особей приводит к расщеплению по цвету оперения среди потомков в соотношении 9 : 3 : 3 : 1.
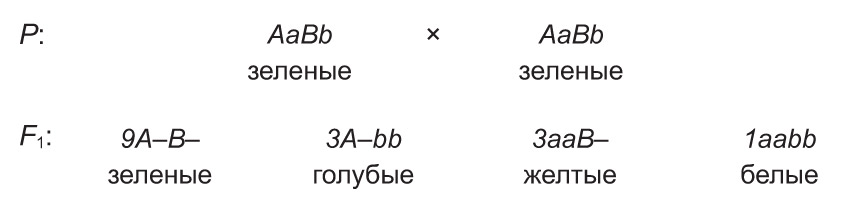
Эпистаз — тип взаимодействия, при котором ген одной аллели подавляет проявление генов другой аллельной пары. Такие гены, блокирующие фенотипическое проявление других неаллельных генов, называются эпистатическими, генами-ингибиторами или супрессорами. Подавляемые гены, в свою очередь, получили название гипостатических. В зависимости от того, доминантным или рецессивным является ген-ингибитор, различают доминантный и рецессивный эпистаз.
Примером доминантного эпистаза может служить взаимодействие двух аллелей, определяющих масть (окраску) лошадей. Так, доминантный ген одной аллельной пары (А) обусловливает вороную (черную) масть лошади, а рецессивный ген (а) — рыжую. Однако эти гены могут проявиться фенотипически только при отсутствии в генотипе лошади доминантного гена другой аллели — ингибитора (I). При наличии ингибитора проявление генов вороной и рыжей масти подавляется. В рассматриваемом примере ген-супрессор имеет собственное фенотипическое проявление: в случае его присутствия у особей развивается серая окраска.
Рассмотрим скрещивание дигетерозиготных лошадей (рис. 33-1.2). Они имеют серую масть, что обусловлено наличием в генотипе доминантного гена-супрессора (I). В потомстве от скрещивания наблюдается расщепление 12 : 3 : 1. Все особи, содержащие в генотипе эпистатический ген (9A—I— и 3aaI—), наследуют серую масть. У жеребят, не имеющих гена I, проявляется вороная либо рыжая окраска в соотношении 3 вороные (3A–ii) к 1 рыжей (1aaii).

Если ген-супрессор является рецессивным, то взаимодействие неаллельных генов протекает по типу рецессивного эпистаза (криптомерии). Такое взаимодействие наблюдается, например, при наследовании цвета семян фасоли. У этого растения красная окраска семян (А) доминирует над желто-коричневой (а). Проявление рецессивного ингибитора (i) ведет к блокировке действия генов красной и желто-коричневой окраски, и семена оказываются белыми. Однако действие рецессивного эпистатического гена может проявиться только при отсутствии в генотипе соответствующего ему доминантного аллеля (I). В случае наличия такого гена действие супрессора подавляется. Так, скрещивание дигетерозиготных растений фасоли приводит к расщеплению среди потомков в соотношении:  с красными,
с красными,  с желто-коричневыми и
с желто-коричневыми и  с белыми семенами.
с белыми семенами.

Эпистаз наблюдается при наследовании многих признаков живых организмов. При различных формах эпистаза и разных особенностях проявления генов расщепление в потомстве от скрещивания дигетерозиготных особей может составлять 13 : 3, 12 : 3 : 1 или 9 : 3 : 4.
У человека описан так называемый «бомбейский феномен». Суть его заключается в том, что у некоторых людей, имеющих в генотипе ген IA или IB, фенотипически проявляется первая (0) группа крови. Было установлено, что в основе этого явления лежит рецессивный эпистаз. Данные люди оказались гомозиготными по редкому рецессивному гену, нарушающему синтез антигенов А и В. Поэтому независимо от сочетания генов, определяющих группу крови по системе АВ0, эритроциты таких людей не содержат антигенов А и В.
Полимерия — взаимодействие, при котором гены двух или более аллелей проявляются сходным образом, определяя развитие одного и того же признака. В случае полимерии неаллельные гены, контролирующие тот или иной признак (полимерные гены), принято обозначать одинаковыми буквами с цифровыми индексами. При этом гены каждой аллельной пары имеют одинаковый индекс. Например: А1, а1 — доминантный и рецессивный аллели первой пары генов, А2, а2 — второй пары и т. д. Известны две разновидности полимерии.
При некумулятивной полимерии доминантный признак проявляется в полной мере при наличии в генотипе хотя бы одного доминантного гена. Иными словами, количество доминантных генов не влияет на степень выраженности признака. Так, у пастушьей сумки форма плодов определяется двумя парами полимерных генов. При этом рецессивный признак — овальные плоды проявляется только у растений, имеющих генотип а1а1а2а2. Если в генотипе присутствует хотя бы один доминантный ген (неважно какой именно — А1 или А2), растение обладает доминантным признаком — плодами треугольной формы.
Скрещивание доминантной и рецессивной дигомозигот (рис. 33-1.3) приводит к появлению в первом поколении дигетерозиготных гибридов с треугольными плодами (А1а1А2а2). Во втором поколении наблюдается расщепление в соотношении 15 : 1.

В случае кумулятивной полимерии степень выраженности доминантного признака определяется количеством доминантных генов в генотипе. Чем больше таких генов, тем сильнее проявляется данный признак. Например, у пшеницы определенных сортов окраска зерен контролируется тремя парами неаллельных генов. Растения с генотипом а1а1а2а2а3а3 имеют белые зерна, а обладающие генотипом А1А1А2А2А3А3 — темно-красные. У растений с другими генотипами наблюдаются различные промежуточные варианты окраски, зависящие от числа доминантных генов. Так, наличие в генотипе только одного доминантного гена обусловливает бледно-розовый цвет зерен, двух — розовый, трех — светло-красный и т. д.
Установлено, что по типу кумулятивной полимерии наследуются многие количественные признаки организмов: рост человека, цвет его кожи (т. е. количество меланина), яйценоскость кур, молочность коров, содержание сахарозы в корнеплодах сахарной свеклы, белков в эндосперме семян злаков и др.
Некоторые гены, не имеющие собственного фенотипического проявления, способны усиливать или, наоборот, ослаблять (в отличие от эпистаза не подавляя полностью) проявление других неаллельных генов. Такие гены называют модификаторами, а их действие — модифицирующим. Например, у кошек черный цвет шерсти доминирует над белым. Гетерозиготные особи обладают черной окраской, но, как правило, один или несколько участков их шерсти имеет белый цвет. Количество и размер таких участков зависят от соответствующих генов-модификаторов. У серых мышей подобным образом определяется размер белого пятна на брюшке. Предполагается, что у человека цвет радужной оболочки глаз контролируется как минимум шестью парами генов. При этом наряду с «основными» генами, определяющими карие и голубые глаза, существенное влияние на окраску радужки оказывают гены-модификаторы. Их действие приводит к появлению разнообразных оттенков — от светло-серого или светло-голубого до практически черного.

Многие признаки живых организмов определяются взаимодействием неаллельных генов. Основными типами такого взаимодействия являются комплементарность, эпистаз и полимерия. При комплементарном действии доминантные гены разных аллелей вместе обусловливают появление нового признака, не определявшегося ни одним из этих генов по отдельности. В случае эпистаза ген одной аллели (эпистатический ген) подавляет проявление генов другой аллельной пары. В зависимости от того, доминантным или рецессивным является эпистатический ген, различают доминантный и рецессивный эпистаз. Полимерия заключается в том, что гены двух или более аллелей проявляются сходно, определяя развитие одного и того же признака. При кумулятивной полимерии степень проявления доминантного признака зависит от числа доминантных генов, при некумулятивной наличие даже одного доминантного гена ведет к полному проявлению доминантного признака.
 |
1. Укажите типы взаимодействия неаллельных генов: Кодоминирование, эпистаз, полимерия, полное доминирование, неполное доминирование, комплементарность. 2. Что представляет собой комплементарное взаимодействие неаллельных генов? Приведите примеры. 3. В чем заключается действие эпистатических генов? Чем доминантный эпистаз отличается от рецессивного? 4. Что такое полимерия? В чем состоит различие между кумулятивной и некумулятивной полимерией? 5. Какие варианты расщепления могут наблюдаться в потомстве от скрещивания дигетерозигот в случае взаимодействия неаллельных генов по типу: а) комплементарности; б) эпистаза; в) некумулятивной полимерии? 6*. У хомяков коричневая окраска шерсти определяется одним доминантным геном, а абрикосовая — другим. Данные гены расположены в разных парах хромосом, их рецессивные аллели приводят к появлению белой окраски. Если же в генотипе хомяка присутствуют оба неаллельных доминантных гена, появляется новый признак — черная окраска. От скрещивания абрикосового самца с коричневой самкой родился детеныш с белой шерстью. Установите генотипы родительских особей. Какое потомство можно ожидать у них в дальнейшем? 7*. Окраска шерсти собак породы лабрадор контролируется двумя аллелями, локализованными в разных парах хромосом. Гены одной аллельной пары обусловливают развитие черной или коричневой окраски, при этом черная масть полностью доминирует над коричневой. В другой аллельной паре рецессивный ген обладает эпистатическим действием. Он подавляет проявление генов черной и коричневой окраски, обусловливая развитие шерсти светло-бежевого цвета. Дигетерозиготную особь скрестили с рецессивной дигомозиготной. Какой фенотип имеют скрещиваемые особи? Какова вероятность появления в их потомстве щенков, имеющих: а) черную; б) коричневую; в) светло-бежевую шерсть? 8*. Допустим, что у растений определенного вида длина лепестков определяется двумя парами полимерных генов, расположенными в негомологичных хромосомах. Особи с длиной лепестков 40 мм имеют полностью доминантный генотип, а с длиной 20 мм — полностью рецессивный. Растения с другими генотипами имеют лепестки длиной 35, 30 или 25 мм. Какова длина лепестков у дигетерозиготных растений? Какое расщепление в потомстве будет наблюдаться при скрещивании этих растений между собой? |