§ 22-1. Темновая фаза фотосинтеза. Значение фотосинтеза. Хемосинтез
Темновая фаза *(на примере С3-фотосинтеза). Как вы знаете, реакции темновой фазы осуществляются в строме хлоропласта независимо от наличия света. Однако для их протекания необходимы такие продукты световой фазы, как АТФ и восстановленный НАДФ.*
Из окружающей среды в хлоропласты поступает углекислый газ. В ходе темновой фазы фотосинтеза происходит его *связывание (фиксация) и* восстановление до органических веществ. *У фотоавтотрофов обнаружено несколько разных способов фиксации СО2 и его последующего использования для синтеза органических соединений. При этом основным, наиболее распространенным механизмом является так называемый С3-путь фотосинтеза. Его можно разделить на несколько этапов.
1. Связывание углекислого газа. В строме хлоропласта молекулу СО2 присоединяет особый акцептор — рибулозо-1,5-дифосфат (РДФ). Это производное пятиуглеродного моносахарида рибулозы. Реакция протекает с использованием воды и катализируется ферментом РДФ-карбоксилазой, на долю которой может приходиться более половины всех белков хлоропластов. Считается, что РДФ-карбоксилаза — самый распространенный белок на Земле. В результате присоединения углекислого газа к РДФ образуется неустойчивое шестиуглеродное соединение, которое распадается на две молекулы фосфоглицериновой кислоты (ФГК):
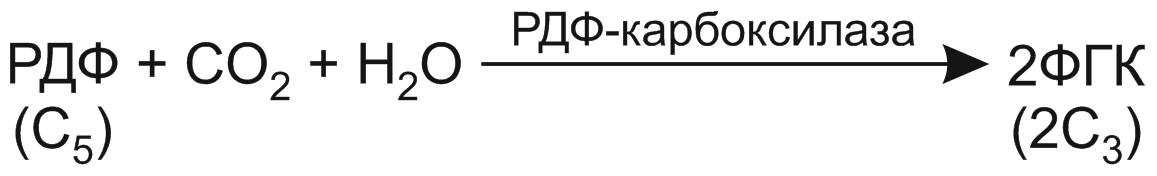
Молекула ФГК — первичного продукта фиксации СО2 — содержит 3 атома углерода. Поэтому данный механизм связывания углекислого газа и назван С3-путем фотосинтеза.
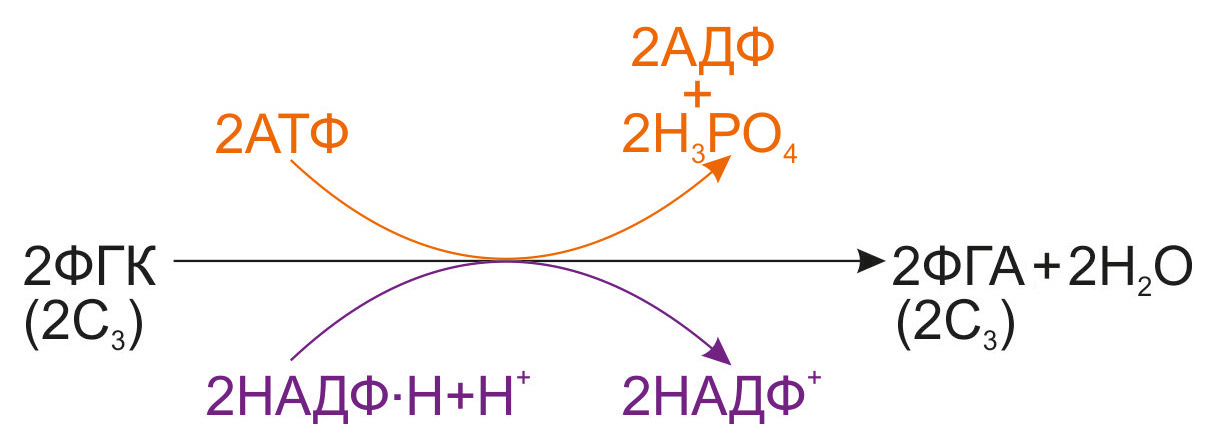
2. Восстановление ФГК. Далее фосфоглицериновая кислота подвергается ферментативному восстановлению до фосфоглицеринового альдегида (ФГА). Процесс протекает с использованием продуктов световой фазы фотосинтеза — АТФ и НАДФ∙Н+Н+:
3. Превращение ФГА в другие продукты фотосинтеза и РДФ. Некоторая часть молекул ФГА идет на синтез глюкозы и других моносахаридов, спиртов, карбоновых кислот, аминокислот (образуются путем аминирования карбоновых кислот) и т. п. Далее из них могут синтезироваться более сложные соединения — олиго- и полисахариды (крахмал, целлюлоза), липиды, белки и др.
Однако бóльшая часть молекул ФГА превращается в рибулозо-1,5-дифосфат, способный снова связывать СО2. Реакции регенерации РДФ протекают с затратами энергии АТФ. Таким образом, С3-путь фиксации углекислого газа представляет собой циклический процесс. В честь американского биохимика М. Кальвина, исследовавшего этот способ ассимиляции СО2 (Нобелевская премия за 1961 г.), он назван циклом Кальвина.*
*За один «оборот» цикла Кальвина фиксируется одна молекула углекислого газа. Поэтому для синтеза шестиуглеродной молекулы глюкозы (С6Н12О6) требуется шесть «оборотов» цикла. Пять из них необходимы для регенерации РДФ, а один идет собственно на образование глюкозы (рис. 22-1.1).*

*При С3-фотосинтезе для образования одной молекулы глюкозы из СО2 нужно использовать, а затем снова регенерировать 6 молекул РДФ, окислить 12 молекул НАДФ∙Н+Н+ (он служит источником атомов водорода) и расщепить 18 молекул АТФ (она является поставщиком энергии). Общее уравнение темновой фазы фотосинтеза можно записать следующим образом (для упрощения не указаны молекулы воды, необходимые для гидролиза АТФ):
6СО2 + 12НАДФ∙Н+Н+ + 18АТФ → С6Н12О6 + 6Н2О + 12НАДФ+ + 18АДФ + 18Н3РО4.*
Следовательно, АТФ *и восстановленный НАДФ,* полученные в ходе световой фазы, используются в темновой фазе для образования глюкозы и других продуктов фотосинтеза. При этом энергия макроэргических связей АТФ преобразуется в энергию химических связей органических веществ.
*Как уже отмечалось, реакции темновой фазы протекают почти одновременно со световой. Эксперименты показали, что на свету в хлоропластах происходит связывание углекислого газа и синтез глюкозы. После выключения света образование глюкозы некоторое время продолжается, а затем останавливается. Но если в среду с хлоропластами добавить АТФ и НАДФ∙Н+Н+, синтез глюкозы возобновится. Он будет протекать в темноте до тех пор, пока не истощатся запасы АТФ и восстановленного НАДФ.*
*Итак, для синтеза молекулы глюкозы необходимо 12 молекул НАДФ∙Н+Н+. Чтобы восстановить 12 молекул НАДФ+ в световой фазе, нужно присоединить к ним 24 атома водорода, т. е. 24 электрона (е–) и 24 протона (Н+). А для их образования требуется подвергнуть фотолизу 12 молекул воды:
12Н2О → 6О2 + 24е– + 24Н+.*
Если объединить процессы, протекающие в световой и темновой фазах, исключив промежуточные продукты *и сократив молекулы воды* (рис. 22-1.2), можно получить суммарное уравнение фотосинтеза:


*Другие пути фиксации СО2 при фотосинтезе. У некоторых растений первичным продуктом фиксации углекислого газа является не ФГК, как при С3-пути фотосинтеза, а четырехуглеродное соединение — щавелевоуксусная кислота (ЩУК). Такой механизм связывания СО2 известен как С4-путь фотосинтеза, или путь Хэтча–Слэка (в честь австралийских ученых М. Д. Хэтча и Ч. Р. Слэка, детально исследовавших этот способ ассимиляции углекислого газа).
Для растений, использующих С4-путь фотосинтеза (например, кукурузы, проса, сорго, сахарного тростника), характерно особое строение листьев. Их проводящие пучки окружены двумя слоями фотосинтезирующих клеток. Внутренний слой образует так называемую обкладку проводящего пучка. Наружный слой представлен клетками мезофилла (основной хлорофиллоносной ткани листьев растений, в большинстве случаев дифференцированной на столбчатую и губчатую паренхиму), в гиалоплазме которых и происходит фиксация СО2 по С4-пути.
Акцептором углекислого газа при С4-фотосинтезе служит трехуглеродное соединение — фосфоенолпировиноградная кислота (ФЕП):
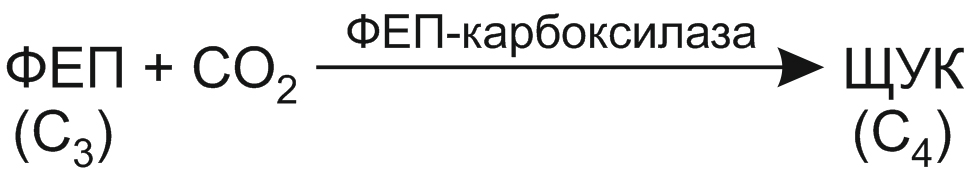
Установлено, что ФЕП-карбоксилаза, катализирующая эту реакцию, более активно связывает СО2, чем РДФ-карбоксилаза, выполняющая такую же функцию в цикле Кальвина.
Щавелевоуксусная кислота (ЩУК), образовавшаяся в клетках мезофилла, превращается в яблочную (или аспарагиновую) кислоту, которая поступает в клетки обкладки проводящего пучка (рис. 22-1.3). Здесь путем отщепления молекулы СО2 она превращается в ПВК. В клетках обкладки осуществляется стандартный С3-путь связывания углекислого газа, т. е. выделившийся СО2 поступает в цикл Кальвина. ПВК возвращается в клетки мезофилла, где превращается в ФЕП — исходный акцептор углекислого газа.*

*Известно, что у С3-растений, фиксирующих СО2 только с помощью цикла Кальвина, фотосинтез протекает быстро лишь при довольно высокой концентрации углекислого газа в воздухе. С4-путь даже в условиях низкого содержания СО2 позволяет накапливать в клетках обкладки углекислый газ в количестве, достаточном для эффективного протекания цикла Кальвина. Следовательно, у С4-растений фотосинтез осуществляется интенсивно и при низких концентрациях СО2 в окружающей среде. При этом отпадает необходимость постоянно держать устьица открытыми (для поступления большего количества СО2 к фотосинтезирующим клеткам), что снижает потери воды в ходе транспирации. Все это позволяет С4-растениям осваивать засушливые, жаркие местообитания.
В подобных условиях могут существовать и растения, использующие так называемый САМ-фотосинтез (сокращение от англ. crassulacean acid metabolism — кислотный метаболизм толстянковых). Этот путь характерен преимущественно для суккулентов — толстянок, кактусов, каланхоэ и др. В связи со строгой экономией воды устьица САМ-растений в дневное время закрыты. Открываются они только ночью, в самый прохладный период суток. Следовательно, углекислый газ может поступать в листья только ночью. В гиалоплазме хлорофиллоносных клеток он фиксируется по С4-пути и накапливается в вакуолях в виде яблочной кислоты (см. рис. 22-1.3). Днем, когда устьица закрыты, яблочная кислота переходит в гиалоплазму и высвобождает запасенный СО2. Он поступает в строму хлоропластов и вовлекается в цикл Кальвина.
Таким образом, механизм САМ-фотосинтеза сходен с С4-фотосинтезом. Различие заключается в том, что у С4-растений связывание углекислого газа по С4- и С3-пути происходит в разных типах клеток, а у САМ-растений — в пределах одной и той же клетки, но в разное время суток.*