§ 18. Мейоз
Мейоз — особый способ деления эукариотических клеток, в результате которого из одной материнской клетки образуются четыре дочерние с уменьшенным в 2 раза набором хромосом. Если в мейоз вступает диплоидная клетка (2n4c), то образуются четыре гаплоидные клетки (1n1c). Клетки с гаплоидным набором хромосом не способны делиться мейозом.
Мейоз представляет собой два последовательных деления — мейоз I и мейоз II. Важно отметить, что репликация ДНК предшествует только первому мейотическому делению. Между мейозом I и мейозом II удвоения ДНК не происходит. Каждое из двух делений обычно включает профазу, метафазу, анафазу и телофазу. Рассмотрим процесс мейотического деления диплоидной клетки. Первое деление мейоза осуществляется следующим образом (табл. 18.1)
Таблица 18.1. Первое деление мейоза (мейоз I)
|
Схема |
Фаза и процессы, происходящие в ней |
|
|
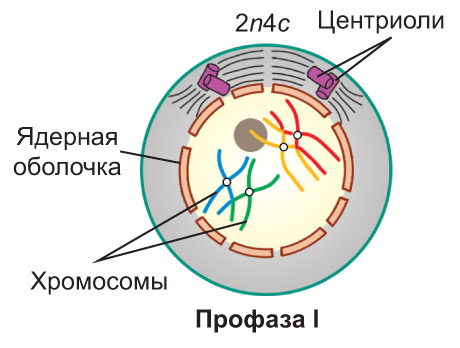
Профаза I. В ядре начинается спирализация хроматина. *Длинные тонкие хроматиновые нити постепенно укорачиваются и при этом утолщаются, и формирующиеся хромосомы становятся четко различимыми в световой микроскоп.* Каждая хромосома состоит из двух хроматид. Содержание наследственного материала в клетке выражается записью 2n4c. Гомологичные хромосомы попарно сближаются и соединяются друг с другом *сначала в нескольких участках, а затем по всей своей длине*. Этот процесс называется конъюгацией гомологичных хромосом, *а образовавшиеся хромосомные пары — бивалентами. В ходе конъюгации хромосомы тесно соприкасаются. В некоторых точках соприкосновения, называемых хиазмами, вследствие разрыва и последующего воссоединения молекул ДНК,* между хроматидами гомологичных хромосом может происходить обмен соответствующими участками — кроссингóвер. *Далее гомологичные хромосомы в составе каждого бивалента начинают отталкиваться друг от друга и в результате остаются связанными только в областях хиазм.* Одновременно с этим в клетке начинается образование веретена деления. К концу профазы I исчезают ядрышки и распадается ядерная оболочка |
|
|
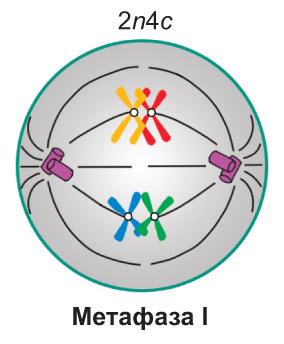
Метафаза I. Завершается формирование веретена деления. Спирализация хромосом достигает максимума. *Биваленты* располагаются в центральной части клетки, *образуя метафазную пластинку. Каждый бивалент ориентирован таким образом, что центромеры гомологичных хромосом находятся по разные стороны от экваториальной плоскости клетки, на одинаковом расстоянии от нее.* При этом нити веретена деления, идущие от противоположных полюсов клетки, прикреплены к центромерам разных гомологичных хромосом. Таким образом, в составе каждого *бивалента* одна из хромосом оказывается связанной с одним полюсом клетки, а другая — с противоположным |
|
|
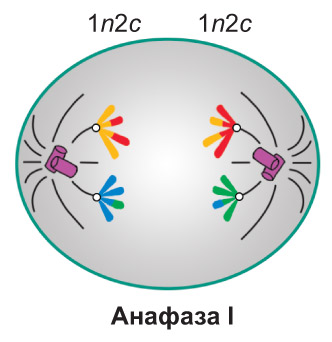
Анафаза I. *Каждый бивалент распадается на две хромосомы, причем их* центромеры, в отличие от митоза, не разделяются надвое. Поэтому нити веретена деления растягивают к противоположным полюсам клетки не сестринские хроматиды, как при митозе, а гомологичные хромосомы. Следовательно, диплоидный набор 2n4c разделяется на два гаплоидных набора, которые впоследствии попадут в разные дочерние клетки. Каждая хромосома состоит из двух хроматид, которые в результате кроссинговера уже не идентичны друг другу. Таким образом, в конце анафазы I набор хромосом и хроматид у каждого полюса делящейся клетки составляет 1n2c |
|
|
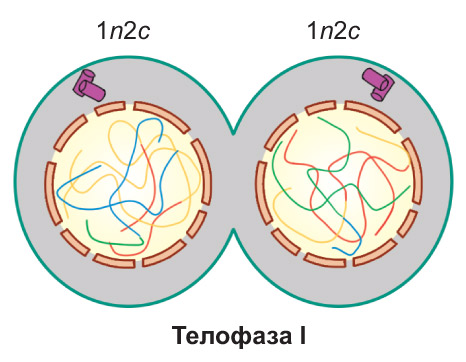
Телофаза I. Веретено деления разрушается. Происходит деспирализация хромосом и формирование двух ядер. Далее клетка разделяется на две дочерние. Они имеют гаплоидный набор хромосом, каждая хромосома состоит из двух хроматид (1n2c) |
Промежуток времени между первым и вторым делениями мейоза обычно очень короткий. В этот период, как уже отмечалось, не осуществляется репликация ДНК. Каждая из двух клеток, образовавшихся в результате мейоза I, вступает в мейоз II. Это деление протекает аналогично митозу (табл. 18.2).
*У некоторых организмов перерыв между мейозом I и мейозом II — так называемый интеркинез вообще отсутствует. При этом клетки, завершившие телофазу I, сразу же переходят в профазу II. Более того, могут выпадать и эти фазы. Так, у большинства растений, клетки, делящиеся мейозом, после окончания анафазы I вступают непосредственно в метафазу II. В этом случае переход от первого мейотического деления ко второму не сопровождается деспирализацией хромосом, формированием двух ядер и разделением материнской клетки на две дочерние.*
Таблица 18.2. Второе деление мейоза (мейоз II)
|
Схема |
Фаза и процессы, происходящие в ней |
|
|
Профаза II. В результате спирализации хроматина формируются двухроматидные хромосомы (набор 1n2c). В это же время начинается образование веретена деления. После распада ядерной оболочки отдельные хромосомы беспорядочно располагаются в гиалоплазме |
|
|
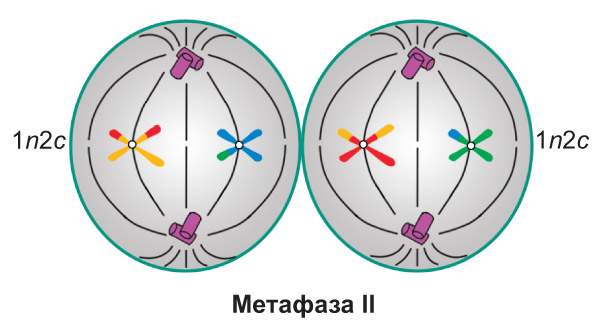
Метафаза II. Завершается формирование веретена деления. Хромосомы, достигшие максимальной спирализации, выстраиваются в центральной части клетки, *формируя метафазную пластинку*. Нити веретена деления связывают центромеру каждой хромосомы с двумя противоположными полюсами |
|
|
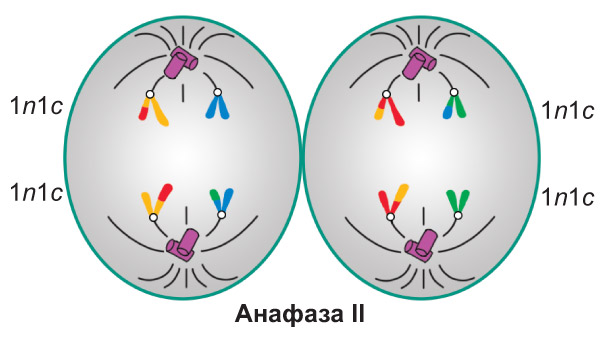
Анафаза II. Происходит разделение центромер. Сестринские хроматиды (теперь уже дочерние хромосомы) растягиваются к разным полюсам клетки. В конце анафазы II набор хромосом и хроматид у каждого полюса составляет 1n1c |
|
|
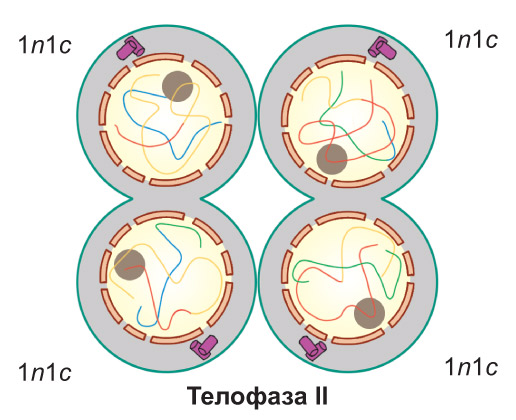
Телофаза II. Хромосомы деспирализуются, формируются ядра, и происходит разделение клеток. При этом образуются четыре дочерние клетки, имеющие набор 1n1c |

Таким образом, в результате первого деления мейоза исходная материнская клетка (2n4c) разделилась на две дочерние, имеющие гаплоидный набор хромосом. *Следовательно, в мейозе I произошла редукция (уменьшение) числа хромосом в 2 раза. Поэтому первое деление называют редукционным. В образовавшихся гаплоидных клетках каждая хромосома представлена двумя хроматидами (1n2c).
В результате второго деления, протекающего по типу митоза, набор хромосом (1n) не изменяется. Это деление мейоза называется эквационным (от лат. aequatio — уравнивание). Эквационное деление, в отличие от редукционного, сопровождается расхождением сестринских хроматид. Поэтому итогом второго деления является образование четырех гаплоидных клеток с однохроматидными хромосомами (1n1c).*
Мейоз — более длительный процесс, чем митоз. Например, у ржи он протекает более 2 сут, у репчатого лука — около 4 сут. Мейотическое деление клеток мыши занимает примерно неделю. Мейоз, протекающий при образовании сперматозоидов человека, длится приблизительно 25 дней. Самой продолжительной стадией мейоза является профаза I.
*Как и в случае митоза, правильное протекание мейоза может быть нарушено действием определенных внешних или внутриклеточных факторов. Последствия таких воздействий могут быть разными, вплоть до образования нежизнеспособных дочерних клеток. Наиболее часто встречающейся патологией мейоза является нерасхождение хромосом в анафазе I. Оно происходит вследствие нарушения разделения бивалентов и выражается в том, что обе гомологичные хромосомы перемещаются к одному из полюсов клетки. Нерасхождение может наблюдаться и на стадии анафазы II. При этом к одному и тому же полюсу отходят две сестринские хроматиды (дочерние хромосомы). Как в первом, так и во втором случае результатом нерасхождения хромосом является образование генетически неравноценных клеток. В одних клетках наблюдается избыток хромосом (1n + 1), а в других — недостаток (1n — 1).*
Биологическое значение мейоза. У животных и человека путем мейоза образуются гаметы — гаплоидные половые клетки. В результате последующего оплодотворения формируется зигота с двойным набором хромосом, из которой развивается новый организм. Он является диплоидным, как и его родители, а значит, сохраняет свойственный данному виду организмов кариотип. Без мейоза, приводящего к уменьшению набора хромосом в 2 раза, половое размножение сопровождалось бы удвоением числа хромосом в каждом новом поколении. У растений, многих водорослей и грибов мейоз приводит к формированию спор, с помощью которых осуществляется бесполое размножение.
Кроссинговер, происходящий в профазе I, приводит к перекомбинации наследственного материала между гомологичными хромосомами. В анафазе I гомологичные хромосомы каждой пары случайным образом, независимо от других пар, расходятся к разным полюсам клетки. В анафазе II то же самое происходит с сестринскими хроматидами. Все эти процессы являются важными источниками комбинативной изменчивости (будет рассмотрена подробно в главе 6), обеспечивающей появление разнообразного потомства как при половом размножении, так и при размножении спорами.