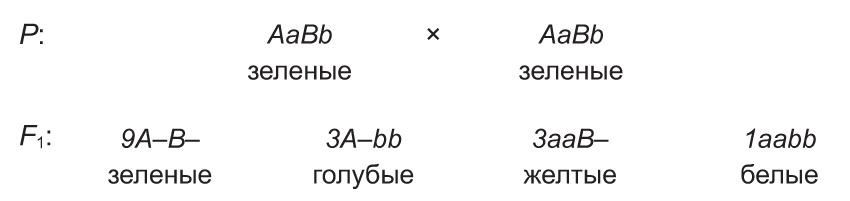*§ 33-1. Узаемадзеянне неалельных генаў
Прыметы, наследаванне якіх мы разглядалі раней, кантралююцца генамі адной алельнай пары. Аднак многія прыметы жывых арганізмаў вызначаюцца ўзаемадзеяннем двух ці больш за пар генаў, г. зн. неалельнымі генамі. Асноўнымі тыпамі ўзаемадзеяння неалельных генаў з'яўляюцца камплементарнасць, эпістаз і полімерыя.
Камплементарнасць выяўляецца ў тым, што дамінантныя гены розных алеляў супольна абумоўліваюць з'яўленне новай прыметы, якая не вызначалася ніводным з гэтых генаў паасобку. Напрыклад, у льну дамінантны ген адной алелі (А) вызначае ружовую афарбоўку кветак, а дамінантны ген другой алелі (В) — белую. Калі ў генатыпе расліны прысутнічаюць абодва дамінантныя алелі, яго кветкі маюць блакітную афарбоўку. Такім чынам, неалельныя гены А і В дзейнічаюць камплементарна (узаемна дапаўняюць адзін аднаго), што прыводзіць да фарміравання новай прыметы. Сутнасць такога ўзаемадзеяння можна адлюстраваць у выглядзе схемы.

Як бачна са схемы, сінтэз блакітнага пігменту з неафарбаванага папярэдніка (прапігменту) ажыццяўляецца ў два этапы, кожны з якіх каталізуецца асаблівым ферментам. Наяўнасць гэтых ферментаў у клетках пялёсткаў лёну вызначаецца наяўнасцю ў генатыпе адпаведных генаў А і В. Так, ген А вызначае наяўнасць ферменту, які ператварае прапігмент у прамежкавы прадукт — пігмент ружовага колеру. Ген В абумоўлівае наяўнасць другога ферменту, які сінтэзуе блакітны пігмент з ружовага. Таму ў раслін, што маюць абодва дамінантныя алелі, у пялёстках утвараецца канчатковы прадукт — пігмент блакітнага колеру. Калі адсутнічае ген В (а значыць, і адпаведны фермент), у клетках працякае толькі першая стадыя — утварэнне ружовага пігменту. У выпадку адсутнасці гена А (ці абодвух дамінантных алеляў) не ажыццяўляецца нават першы этап сінтэзу пігменту, і пялёсткі застаюцца белымі.
Разгледзім скрыжаванне дыгомазіготных раслін лёну, якія маюць ружовыя (AAbb) і белыя (aaBB) кветкі (мал. 33-1.1). У першым пакаленні назіраецца аднастайнасць гібрыдаў. Усе яны маюць блакітныя кветкі, што абумоўлена наяўнасцю ў генатыпе як гена А, так і гена В. У другім гібрыдным пакаленні назіраецца расшчапленне ў суадносінах 9 блакітных : 3 ружовых : 4 белых.

Камплементарнае ўзаемадзеянне генаў ляжыць у аснове праяўлення шэрага прымет у розных арганізмаў. Так, па тыпе камплементарнасці ўзаемадзейнічаюць гены, што кантралююць афарбоўку коканаў у тутавага шаўкапрада і кветак у духмянага гарошку, форму грэбеня ў курэй, колер апярэння ў папугаяў і г. д. У залежнасці ад асаблівасцей фенатыпічнага праяўлення генаў у патомства ад скрыжавання дыгетэразігот могуць назірацца розныя варыянты расшчаплення — 9 : 7, 9 : 3 : 4, 9 : 6 : 1, 9 : 3 : 3 : 1.
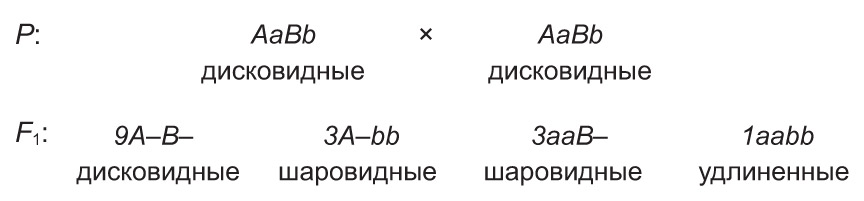
Напрыклад, у гарбуза дамінантныя алелі А і В супольна прыводзяць да з'яўлення пладоў дыскападобнай формы, а паасобку — шарападобнай. У той жа час рэцэсіўныя алелі гэтых генаў абумоўліваюць падоўжаную форму пладоў. Пры скрыжаванні дыгетэразіготных раслін, якія маюць дыскападобныя плады, у патомстве адбываецца расшчапленне 9 : 6 : 1.
Падобным чынам наследуецца колер апярэння хвалістых папугаяў. Дамінантны ген адной алелі (А) абумоўлівае ў іх блакітную афарбоўку пёраў, а дамінантны ген другой алелі (В) — жоўтую. Пры наяўнасці ў генатыпе абодвух дамінантных алеляў (А і В) выяўляецца новая прымета — зялёнае апярэнне. Разам з тым рэцэсіўныя алелі гэтых генаў (a і b) вызначаюць белую афарбоўку пёраў. Таму скрыжаванне зялёных дыгетэразіготных асобін прыводзіць да расшчаплення па колеры апярэння сярод нашчадкаў у суадносінах 9 : 3 : 3 : 1.