§ 17. Простае бінарнае дзяленне. Мітоз. Амітоз
| Сайт: | Профильное обучение |
| Курс: | Біялогія. 11 клас |
| Книга: | § 17. Простае бінарнае дзяленне. Мітоз. Амітоз |
| Напечатано:: | Гость |
| Дата: | Воскресенье, 6 Июль 2025, 08:29 |
Простае бінарнае дзяленне. Клеткі пракарыётаў у пераважнай большасці выпадкаў дзеляцца з утварэннем двух аднолькавых па памеры даччыных клетак. Гэты працэс называецца *простым бінарным дзяленнем* (простым дзяленнем папалам).
*Перад дзяленнем адбываецца рэплікацыя бактэрыяльнай храмасомы — кальцавой малекулы ДНК, якая ў пэўным пункце прымацавана да плазмалемы. Пры гэтым утвараюцца дзве ідэнтычныя даччыныя храмасомы, таксама прымацаваныя да цытаплазматычнай мембраны (мал. 17.1). Потым спецыяльныя маторныя бялкі перамяшчаюць даччыныя храмасомы ў процілеглых напрамках, і адлегласць паміж малекуламі ДНК павялічваецца.

У працэсе дзялення плазмалема ўгінаецца (урастае) унутр клеткі паміж дзвюма храмасомамі, якія разышліся. Адначасова ў гэтай вобласці адбываецца дабудаванне клетачных сценак будучых даччыных клетак. У канчатковым выніку бактэрыяльная клетка дзеліцца папалам. Пры гэтым у кожнай даччынай клетцы аказваецца па адной храмасоме. Простае бінарнае дзяленне ўласцівае толькі пракарыётам.* Эўкарыятычныя клеткі, у адрозненне ад пракарыятычных, маюць больш складаную арганізацыю. Таму для іх характэрны іншыя спосабы дзялення: мітоз, амітоз, меёз.
Мітоз — асноўны спосаб дзялення клетак эўкарыёт, у выніку якога з адной мацярынскай клеткі ўтвараюцца дзве даччыныя з такім жа наборам храмасом, як і ў мацярынскай клетцы. Працягласць мітозу ў сярэднім складае 1—2 гадзіны. Пры гэтым клеткі жывёл, як правіла, дзеляцца хутчэй (30—60 мін), чым раслінныя (2—3 г). Мітозам могуць дзяліцца клеткі з рознымі наборамі храмасом — гаплоідныя (1n), дыплоідныя (2n), трыплоідныя (3n) і г. д. Разгледзім працэс мітатычнага дзялення на прыкладзе дыплоіднай клеткі (табл. 17.1).
Табліца 17.1. Мітоз
|
Схема |
Фаза і працэсы, якія ў ёй адбываюцца |
|
|
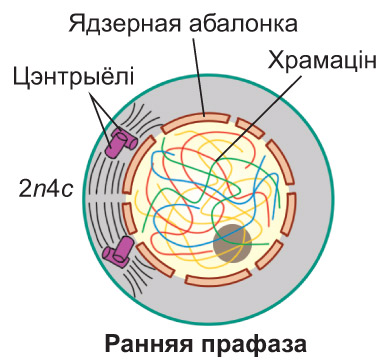
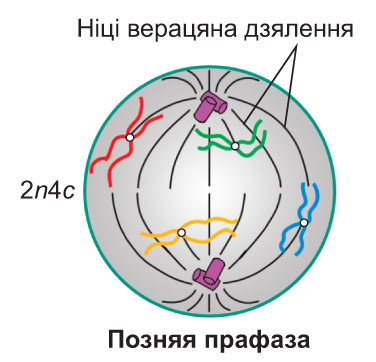
Прафаза. У ядры клеткі пачынаецца спіралізацыя храмаціну, што паступова прыводзіць да фарміравання храмасом. *У пачатку прафазы ніці храмаціну, сабраныя ў клубок, ледзь бачны ў светлавы мікраскоп. Паступова яны ўкарочваюцца і адначасова патаўшчаюцца, так што да канца прафазы ўжо можна выразна бачыць асобныя храмасомы.* Кожная з іх складаецца з дзвюх сястрынскіх храматыд, злучаных у вобласці цэнтрамеры. *У гэтым участку храмасомы размяшчаюцца асаблівыя бялковыя структуры — кінетахоры, да якіх пасля будуць прымацоўвацца ніці верацяна дзялення. Як правіла, кожная храматыда мае па адным кінетахоры. Падчас фарміравання храмасом знікаюць ядзеркі. Абалонка ядра распадаецца на дробныя фрагменты. Часткова спіралізаваныя храмасомы аказваюцца ў гіялаплазме і размяшчаюцца ў ёй хаатычна. Набор храмасом і храматыд у клетцы можна адлюстраваць запісам 2n4c. У час прафазы два клетачныя цэнтры (падваенне гэтага арганоіда, як вы ведаеце, адбылося ў S-перыядзе інтэрфазы) ініцыююць утварэнне мікратрубачак. З іх пачынае фарміравацца верацяно дзялення. У працэсе яго ўтварэння цэнтрыёлі парамі разыходзяцца да процілеглых полюсаў клеткі. *Частка ніцей верацяна дзялення накіроўваецца ад аднаго полюса да процілеглага. Іншыя ніці прымацоўваюцца да кінетахораў і спрыяюць перамяшчэнню храмасом у экватарыяльную плоскасць клеткі. Акрамя таго, фарміруюцца кароткія мікратрубачкі, якія адыходзяць ад полюсаў клеткі радыяльна, утвараючы так званую фігуру зоркі.* У клетках, якія не маюць клетачная цэнтра (што характэрна, напрыклад, для большасці раслін), верацяно дзялення фарміруецца без удзелу цэнтрыёлей. *У гэтым выпадку цэнтрамі арганізацыі мікратрубачак з'яўляюцца бесструктурныя аморфныя зоны, размешчаныя на полюсах клеткі. Іх называюць полярнымі шапачкамі* |
|
|
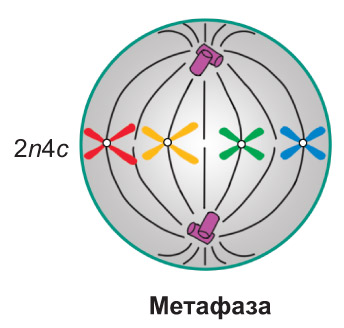
Метафаза. Завяршаецца фарміраванне верацяна дзялення. Храмасомы дасягаюць максімальнай спіралізацыі і размяшчаюцца ў цэнтральнай частцы клеткі, прыкладна на роўнай адлегласці ад полюсаў, *утвараючы так званую метафазную пласцінку*. Пры гэтым іх цэнтрамеры знаходзяцца ў экватарыяльнай плоскасці клеткі, *а кінетахоры сястрынскіх храматыд, звязаныя з ніцямі верацяна дзялення, накіраваны да двух процілеглых полюсаў*. У метафазе спіралізацыя храмасом дасягае максімуму. *Менавіта ў гэты перыяд найбольш зручна вывучаць асаблівасці будовы храмасом і падлічваць іх агульную колькасць у клетцы* |
|
|
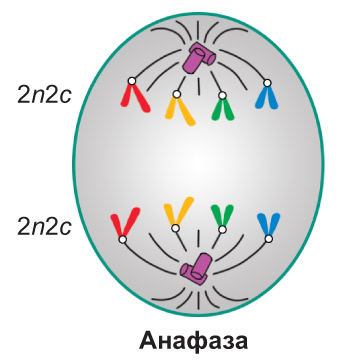
Анафаза. Цэнтрамера кожнай храмасомы дзеліцца папалам, і сястрынскія храматыды аддзяляюцца адна ад адной. З гэтага моманту іх называюць даччынымі храмасомамі. Ніці верацяна дзялення, прымацаваныя да цэнтрамер, скарачаюцца, і даччыныя храмасомы разыходзяцца да процілеглых полюсаў клеткі. *Як правіла, пры гэтым яны прымаюць V-падобную форму: цэнтрамеры накіраваны да полюсаў, а плечы — да экватарыяльнай плоскасці клеткі. Перамяшчэнне ўсіх даччыных храмасом да полюсаў адбываецца з аднолькавай хуткасцю.* У канцы анафазы каля кожнага полюса клеткі аказваецца ідэнтычны набор даччыных храмасом (малекул ДНК) — 2n2c. *У клетках, якія не маюць клетачнай сценкі (у жывёл і інш.), анафаза суправаджаецца расцяжэннем клеткі ў даўжыню. Для раслінных і іншых клетак, пакрытых клетачнай сценкай, такая з'ява не характэрная* |
|
|
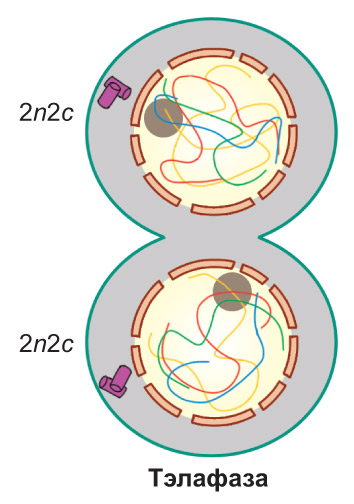
Тэлафаза. Ніці верацяна дзялення паступова разбураюцца. Паблізу ад кожнага полюса клеткі адбываецца дэспіралізацыя (раскручванне) даччыных храмасом з утварэннем храмаціну. Адначасова з гэтым вакол храмасом, якія дэспіралізуюцца, фарміруюцца абалонкі двух новых ядраў. Далей ва ўтвораных ядрах узнікаюць ядзеркі. *На гэтым завяршаецца дзяленне ядра — карыякінез — і пачынаецца дзяленне цытаплазмы з утварэннем дзвюх даччыных клетак — цытакінэз. Перад цытакінэзам арганоіды клеткі, як правіла, больш-менш раўнамерна размяркоўваюцца па цытаплазме. *У экватарыяльнай плоскасці клетак жывёл кампаненты цыташкілета фарміруюць кальцавую перацяжку. Яна паглыбляецца, пакуль не адбудзецца поўны падзел дзвюх даччыных клетак. Клеткі раслін у сувязі з наяўнасцю цвёрдай клетачнай сценкі падзяляюцца інакш. *У экватарыяльную плоскасць расліннай клеткі накіроўваюцца пузыркі комплексу Гольджы. Тут яны зліваюцца адзін з адным. Пры гэтым змесціва пузыркоў выкарыстоўваецца для пабудовы так званай пасярэдняй пласцінкі, а з мембран пузыркоў па абодва бакі ад яе дабудоўваюцца цытаплазматычныя мембраны будучых даччыных клетак. Пасярэдняя пласцінка служыць асновай для фарміравання клетачных сценак новых клетак. У асобных яе ўчастках ёсць поры. Дзякуючы гэтаму паміж утворанымі даччынымі клеткамі захоўваюцца цяжы цытаплазмы (цытаплазматычныя мосцікі), якія, як вы ўжо ведаеце, называюцца плазмадэсмамі. У канчатковым выніку сярэдняя пласцінка злучаецца з клетачнай сценкай мацярынскай клеткі, і мацярынская клетка аказваецца падзеленай на дзве даччыныя* |
Запомніць паслядоўнасць фаз мітатычнага дзялення клеткі дапаможа слова «ПрыМАТ».
Як ужо адзначалася, пры мітозе ўтвараюцца дзве даччыныя клеткі з аднолькавым наборам храмасом — гэткім жа, як у зыходнай мацярынскай клетцы, якая ўступала ў мітоз. Адрозненне заключаецца толькі ў тым, што кожная храмасома даччынай клеткі, якая ўступае ў інтэрфазу новага клетачнага цыкла, да рэплікацыі прадстаўлена не дзвюма ідэнтычнымі храматыдамі, а адной.
Біялагічнае значэнне мітозу. У ходзе мітозу малекулы ДНК, якія змяшчаліся ў ядры мацярынскай клеткі, дакладна і раўнамерна размяркоўваюцца паміж даччынымі. Значыць, дзве новыя клеткі атрымліваюць аднолькавую спадчынную інфармацыю і аказваюцца генетычна ідэнтычнымі адна адной і мацярынскай клетцы. Такім чынам, мітатычнае дзяленне забяспечвае дакладную перадачу генетычнай інфармацыі ў шэрагу пакаленняў клетак і абумоўлівае падтрыманне пастаяннага ліку храмасом.
Дзякуючы мітозу ў мнагаклетачным арганізме адбываецца павелічэнне колькасці клетак. Гэта ляжыць у аснове росту і развіцця ўсіх мнагаклетачных арганізмаў, а таксама забяспечвае працэсы рэгенерацыі — аднаўлення пашкоджаных тканак і органаў. Бясполае размнажэнне многіх арганізмаў (дзяленне аднаклетачных пратыстаў, пачкаванне кішачнаполасцевых, вегетатыўнае размнажэнне раслін і г. д.) таксама абумоўлена мітатычным дзяленнем клетак.
*Нармальны ход мітозу можа быць парушаны рознымі знешнімі ці ўнутрыклетачнымі фактарамі. Напрыклад, дзеянне іянізуючых выпраменьванняў і пэўных хімічных рэчываў можа прыводзіць да разрыву храмасом на асобныя фрагменты. Пры гэтым некаторыя з іх аказваюцца пазбаўленымі цэнтрамеры. Такія ўчасткі храмасом не могуць звязвацца з ніцямі верацяна дзялення і, такім чынам, перамяшчацца пад іх дзеяннем. Калі ў час тэлафазы храмасомны фрагмент, пазбаўлены цэнтрамеры, знаходзіцца каля аднаго з полюсаў, ён можа быць уключаны ў састаў аднаго з даччыных ядзер. Калі ж падобны ўчастак храмасомы размяшчаецца паблізу экватарыяльнай плоскасці клеткі, імавернасць яго ўваходжання ў ядро адной з клетак, якія фарміруюцца, практычна роўна нулю. Такія храмасомныя фрагменты ў далейшым расшчапляюцца.
Часам з прычыны няправільнага фарміравання ніцей верацяна дзялення сястрынскія храматыды той ці іншай храмасомы, якія аддзяліліся адна ад адной, перамяшчаюцца да аднаго і таго ж полюса. У выніку адна з даччыных клетак атрымлівае лішнюю храмасому (2n + 1), а ў другоі клетцы, наадварот, адна з храмасом не будзе мець парнай, гамалагічнай храмасомы (2n — 1). Дзеянне на клетку некаторых хімічных рэчываў можа прыводзіць да змянення вязкасці гіялаплазмы ці парушэння працэсаў зборкі і распаду мікратрубачак верацяна дзялення. Пры гэтым у анафазе часта назіраецца несінхроннае разыходжанне даччыных храмасом да полюсаў. Адны з іх рухаюцца хутчэй, а другія — павольней. У выніку «спозненыя» храмасомы не ўключаюцца ў састаў даччыных ядзер, а ўтвараюць у цытаплазме так званыя мікраядры, якія пасля разбураюцца.
Вядомы хімічныя злучэнні, якія разбураюць ніці верацяна дзялення, але не ўплываюць на працэс падзелу сястрынскіх храматыд. Адным з такіх рэчываў з'яўляецца алкалоід калхіцын. Яго ўплыў на клетку, якая знаходзіцца ў метафазе мітозу, прыводзіць да распаду мікратрубачак верацяна дзялення. Далей сястрынскія храматыды дзеляцца, але без удзелу верацяна дзялення яны не могуць разысціся да полюсаў і застаюцца ў цэнтральнай частцы клеткі. Праз пэўны час вакол гэтых храматыд (зараз ужо даччыных храмасом) утвараецца агульная ядзерная абалонка, і яны ўваходзяць у састаў аднаго ядра. Так узнікае поліплоідная клетка, якая змяшчае набор храмасом, падвоены ў параўнанні з зыходным.
Сустракаюцца таксама выпадкі, калі ў тэлафазе пасля ўтварэння двух ядзер не адбываецца цытакінэз. З прычыны гэтага клетка аказваецца двух'ядзернай.
Такім чынам, парушэнне правільнага працякання мітозу можа прыводзіць да нераўнамернага размеркавання генетычнага матэрыялу паміж даччынымі клеткамі, узнікнення поліплоідных ці двух'ядзерных клетак і г. д.*
Амітоз. Пры амітозе ў клетцы не фарміруецца верацяно дзялення, у яе ядры не адбываецца спіралізацыя храмаціну, захоўваюцца ядзеркі і ядзерная абалонка. Ядро застаецца ў стане, характэрным для інтэрфазы, і падзяляецца папалам перацяжкай. Пры гэтым малекулы ДНК (храмасомы) размяркоўваюцца паміж даччынымі ядрамі нераўнамерна, выпадковым чынам. У шэрагу выпадкаў амітатычнае дзяленне ядра не суправаджаецца наступным дзяленнем клеткі на дзве даччыныя *цытакінэзам*. Гэта прыводзіць да з’яўлення двух’ядзерных і нават шмат’ядзерных клетак. Калі ж даччыныя клеткі ўсё-такі ўтвараюцца, то клетачныя кампаненты, як і ДНК, размяркоўваюцца паміж імі выпадкова і нераўнамерна.
Амітоз — параўнальна рэдкая з’ява. Гэтым спосабам дзеліцца, напрыклад, вялікае ядро інфузорый. У мнагаклетачных арганізмаў амітоз назіраецца пры розных паталагічных працэсах (рост пухлін, запаленне і інш.), а таксама ў клетках, якія старэюць ці павінны загінуць. Як правіла, клеткі, што ўзніклі ў выніку амітозу, губляюць здольнасць уступаць у звычайны клетачны цыкл і ў далейшым дзяліцца мітозам.

Для пракарыятычных клетак характэрна простае бінарнае дзяленне, для эўкарыятычных — мітоз, амітоз і меёз. Мітоз — асноўны спосаб дзялення клетак эўкарыёт, у выніку якога з адной мацярынскай клеткі ўтвараюцца дзве даччыныя з такім жа наборам храмасом. У ходзе мітозу адбываецца спіралізацыя храмаціну з утварэннем кампактных храмасом і фарміраванне верацяна дзялення. Потым сястрынскія храматыды кожнай храмасомы аддзяляюцца адна ад адной і з дапамогай верацяна дзялення раўнамерна размяркоўваюцца паміж утворанымі даччынымі клеткамі. У мнагаклетачных арганізмаў мітатычнае дзяленне клетак забяспечвае працэсы росту, развіцця і рэгенерацыі. У аснове бясполага размнажэння многіх арганізмаў таксама ляжыць мітоз.
Пры амітозе ядро клеткі дзеліцца перацяжкай без спіралізацыі храмаціну і ўтварэння верацяна дзялення. Гэта прыводзіць да нераўнамернага размеркавання малекул ДНК паміж даччынымі ядрамі. У многіх выпадках амітатычнае дзяленне ядра не суправаджаецца наступным дзяленнем клеткі на дзве даччыныя.
 |
1. Якія спосабы дзялення характэрныя для клетак пракарыёт? Для эўкарыятычных клетак? Амітоз, мітоз, меёз, простае дзяленне папалам. 2. Што такое мітоз? Ахарактарызуйце фазы мітозу. 3. У сувязі з чым даччыныя клеткі, утвораныя ў выніку мітозу, атрымліваюць аднолькавую спадчынную інфармацыю? У чым заключаецца біялагічнае значэнне мітозу? 4. Устанавіце адпаведнасць паміж саматычнымі клеткамі чалавека, якія знаходзяцца ў розных перыядах інтэрфазы і мітозу, і колькасцю храмасом і храматыд у гэтых клетках.
5*. У чым заключаюцца адрозненні паміж мітозам і амітозам? Як вы думаеце, чаму мітоз называюць непрамым дзяленнем клеткі, а амітоз — прамым? 6*. У ядры клеткі, якая не дзеліцца, спадчынны матэрыял (ДНК) знаходзіцца ў выглядзе аморфнага разасяроджанага рэчыва — храмаціну. Перад дзяленнем храмацін спіралізуецца і ўтварае кампактныя структуры — храмасомы, а пасля дзялення вяртаецца ў зыходны стан. Для чаго клеткі ажыццяўляюць такія складаныя відазмяненні свайго спадчыннага матэрыялу? |