§ 37. Генатыпічная зменлівасць
| Сайт: | Профильное обучение |
| Курс: | Біялогія. 11 клас |
| Книга: | § 37. Генатыпічная зменлівасць |
| Напечатано:: | Гость |
| Дата: | Понедельник, 18 Август 2025, 22:15 |
Вылучаюць два віды генатыпічнай (спадчыннай) зменлівасці: камбінатыўную і мутацыйную.
Камбінатыўная зменлівасць. З курса біялогіі 10-га класа вам вядома, што для палавога размнажэння характэрна з’яўленне ў бацькоўскіх асобін разнастайнага патомства. Арганізмы новага пакалення адрозніваюцца як ад бацькоўскіх форм, так і адзін ад аднаго. Галоўная прычына гэтых адрозненняў заключаецца ў тым, што пры палавым размнажэнні кожны нашчадак наследуе ўнікальнае спалучэнне генаў сваіх бацькоў. Зменлівасць, абумоўленую ўзнікненнем у патомства новых спалучэнняў (камбінацый) бацькоўскіх генаў, называюць камбінатыўнай. Структура саміх генаў пры гэтым не змяняецца.
Крыніцамі камбінатыўнай зменлівасці з’яўляюцца наступныя працэсы.
● Кросінговер, які адбываецца ў прафазе I меёзу.
● Незалежнае разыходжанне гамалагічных храмасом у анафазе I меёзу і сястрынскіх храматыд (даччыных храмасом) у анафазе II.
● Выпадковае зліццё гамет пры апладненні.
Першыя два працэсы забяспечваюць фарміраванне гамет з рознымі камбінацыямі генаў. Выпадковае зліццё палавых клетак прыводзіць да ўтварэння зігот з рознымі спалучэннямі генаў абодвух бацькоў. У выніку ў патомства з’яўляюцца новыя камбінацыі бацькоўскіх прымет, а таксама новыя прыметы, якіх не было ў бацькоў.
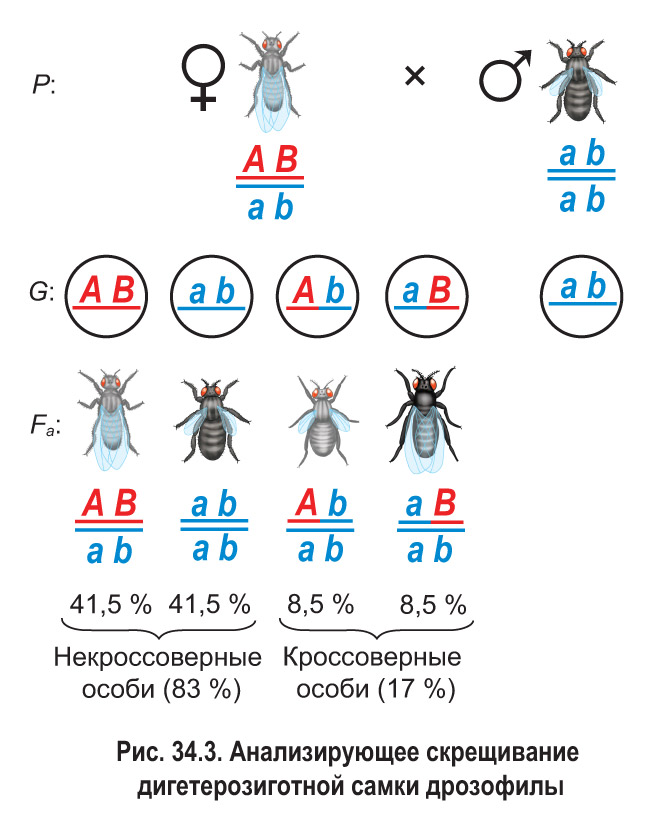
Прыкладамі камбінатыўнай зменлівасці могуць служыць нараджэнне дзяцей з кроўю I ці IV групы ў гетэразіготных бацькоў, якія маюць кроў II і III груп, ці блакітнавокага дзіцяці ў каравокіх гетэразіготных бацькі і маці. Пры гэтым у нашчадкаў узнікаюць новыя прыметы, адрозныя ад бацькоўскіх. Яшчэ адным прыкладам можа быць з’яўленне мух з шэрым целам і зачаткавымі крыламі, з чорным целам і нармальнымі крыламі пры скрыжаванні дыгетэразіготнай дразафілы (шэрае цела, нармальныя крылы) з чорным самцом з зачаткавымі крыламі (гл. мал. 34.3). У дадзеным выпадку ў патомства ў выніку кросінговера з’яўляюцца новыя спалучэнні прымет бацькоў.
{kind=link}
Такім чынам, камбінатыўная зменлівасць з’яўляецца важным пастаўшчыком разнастайнасці жывых арганізмаў.
Мутацыйная зменлівасць. Прычынай гэтага тыпу зменлівасці з’яўляюцца мутацыі — наследуемыя змяненні генетычнага матэрыялу арганізмаў: структуры асобных генаў, будовы ці колькасці храмасом. Працэс узнікнення мутацый называецца мутагенезам, а арганізмы, якія змянілі фенатып у выніку мутацыі, — мутантамі.
Тэрмін «мутацыя» ўпершыню быў прапанаваны нідэрландскім вучоным Х. дэ Фрызам. Ён ахарактарызаваў найважнейшыя ўласцівасці мутацый у працы, якая выйшла ў 1903 г. пад назвай «Мутацыйная тэорыя». Справядлівасць асноўных палажэнняў тэорыі дэ Фрыза была пасля пацверджана шматлікімі даследаваннямі генетыкаў.
Паводле сучасных уяўленняў мутацыі (у адрозненне ад мадыфікацый) не развіваюцца паступова, а ўзнікаюць раптоўна. Яны не ўтвараюць неперарыўных радоў зменлівасці і не маюць нормы рэакцыі. Мутацыі не носяць масавы характар, а праяўляюцца індывідуальна. Найважнейшай уласцівасцю мутацый з’яўляецца іх нявызначанасць. Гэта значыць, што пры мутагенезе можа змяніцца любы элемент спадчыннага матэрыялу клеткі (ген, храмасома, храмасомны набор), прычым нельга дакладна прадказаць, якія менавіта генетычныя структуры будуць закрануты, якім чынам адбудуцца змяненні і да якіх наступстваў гэта прывядзе. Паколькі мутацыі суправаджаюцца змяненнем генатыпу арганізмаў, яны здольныя перадавацца па спадчыне.
Фактары, якія выклікаюць з’яўленне мутацый, называюць мутагеннымі фактарамі ці мутагенамі. У залежнасці ад прыроды адрозніваюць фізічныя, хімічныя і біялагічныя мутагены. Фізічнымі мутагенамі з’яўляюцца розныя віды выпраменьванняў (гама-, рэнтгенаўскае, ультрафіялетавае і інш.), вельмі высокая ці нізкая тэмпература і г. д. Да хімічных мутагенаў належаць разнастайныя рэчывы, напрыклад калхіцын, фармальдэгід, нітрыты, кампаненты тытунёвага дыму, некаторыя харчовыя дабаўкі, пестыцыды і лекавыя прэпараты. Прыкладамі біялагічных мутагенаў могуць быць вірусы.
 Існуюць розныя падыходы да класіфікацыі мутацый. Разгледзім некаторыя з іх.
Існуюць розныя падыходы да класіфікацыі мутацый. Разгледзім некаторыя з іх.
● Паводле тыпу клетак, якія муціравалі, вылучаюць генератыўныя і саматычныя мутацыі. Генератыўнымі называюць мутацыі, якія ўзнікаюць у палавых клетках. Гэтыя мутацыі перадаюцца патомству пры палавым размнажэнні. Саматычныя мутацыі адбываюцца ў саматычных клетках. Такія мутацыі могуць перадавацца па спадчыне шляхам вегетатыўнага размнажэння і праяўляцца ў самой асобіны-мутанта. Пры дзяленні клеткі, якая муціравала, саматычная мутацыя перадаецца даччыным клеткам. Таму, чым раней у ходзе індывідуальнага развіцця ўзнікае такая мутацыя, тым большую частку арганізма яна закранае. Вынікамі праяўлення саматычных мутацый могуць быць, напрыклад, наяўнасць пасмаў валасоў без меланіну ці розны колер вачэй у чалавека (мал. 37.1), з’яўленне парастка з белымі ягадамі на кусце чорнай парэчкі і г. д.
● *У залежнасці ад прычын, якія выклікаюць мутацыі, іх падзяляюць на спантанныя і індуцыраваныя. Спантанныя мутацыі ўзнікаюць у жывых арганізмаў самаадвольна, у звычайных для іх умовах асяроддзя, натуральным шляхам, г. з. без мэтанакіраванага ўмяшання чалавека. Індуцыраванымі называюць мутацыі, штучна выкліканыя з дапамогай мутагенных фактараў у эксперыментальных умовах. Такія мутацыі ўзнікаюць у шмат разоў часцей, чым спантанныя. Іх атрыманне мае важнае значэнне як для вывучэння мутацыйнага працэсу, так і для селекцыйнай працы. Індуцыраваны мутагенез дазваляе павышаць генетычную разнастайнасць арганізмаў, ствараючы тым самым матэрыял для штучнага адбору. На аснове мутантаў, атрыманых такім шляхам, быў створаны шэраг каштоўных штамаў мікраарганізмаў і сартоў раслін.*
 ● Паводле ўзроўню змянення генетычнага матэрыялу мутацыі бываюць геннымі, храмасомнымі і геномнымі.
● Паводле ўзроўню змянення генетычнага матэрыялу мутацыі бываюць геннымі, храмасомнымі і геномнымі.
Генныя мутацыі — гэта змяненні нуклеатыднай паслядоўнасці ДНК у межах аднаго гена з прычыны замены, выпадзення ці ўстаўкі нуклеатыдаў. Генныя мутацыі — самы распаўсюджаны тып мутацый і крыніца з’яўлення новых алеляў. *Такія мутацыі могуць узнікаць не толькі пад дзеяннем мутагенаў, але і, напрыклад, у выніку памылак, якія адбываюцца пры рэплікацыі ДНК.* Змяненні парадку нуклеатыдаў у саставе генаў аднаўляюцца ў структуры адпаведных мРНК і ў большасці выпадкаў вядуць да змянення амінакіслотнай паслядоўнасці бялкоў, якія кадзіруюцца дадзенымі генамі.
Так, у чалавека замена пэўнага нуклеатыду ў гене, які кадзіруе ланцуг гемаглабіну, прыводзіць да замены ў гэтым бялку адной амінакіслаты (глутамінавай) на другую (валін). Змяненне структуры гемаглабіну вядзе да таго, што эрытрацыты замест дыскападобнай формы набываюць серпападобную і губляюць здольнасць да транспарту кіслароду (мал. 37.2). Гэта захворванне называецца серпападобнаклетачнай анеміяй.
Ген, які вызначае форму эрытрацытаў у хворых на серпападобнаклетачную анемію, з’яўляецца рэцэсіўным. Ён кадзіруе асаблівы, анамальны тып гемаглабіну — так званы гемаглабін S. У гетэразіготных носьбітаў гена серпападобнаклетачнай анеміі ў эрытрацытах прысутнічае як звычайны гемаглабін, так і гемаглабін S. Захворванне пры гэтым не развіваецца. Цікава тое, што гемаглабін S абумоўлівае ўстойлівасць эрытрацытаў да заражэння малярыйным плазмодыем. Таму ў рэгіёнах Зямлі, дзе распаўсюджана малярыя, гетэразіготныя носьбіты дэфектнага гена маюць пэўную перавагу перад гомазіготнымі людзьмі. У адрозненне ад рэцэсіўных гомазіготных людзей яны не хварэюць на серпападобнаклетачную анемію і адначасова з’яўляюцца неўспрымальнымі да малярыі, якой могуць хварэць людзі, гомазіготныя па дамінантным гене.
Храмасомныя мутацыі — гэта змяненні будовы храмасом. *Яны ўзнікаюць у выніку разрыву храмасом на фрагменты і наступнага злучэння гэтых фрагментаў у новых спалучэннях.* Змяненні звычайнай структуры адной храмасомы (ці двух гамалагічных) адносяць да ўнутрыхрамасомных мутацый. Перабудовы, у якія былі ўцягнуты негамалагічныя храмасомы, называюць міжхрамасомнымі мутацыямі.
Прыкладамі ўнутрыхрамасомных мутацый могуць быць: выпадзенне ўчастка храмасомы — дэлецыя, двухразовы ці шматразовы паўтор фрагмента храмасомы — дуплікацыя, паварот участка храмасомы на 180° — інверсія. Да міжхрамасомных мутацый належыць абмен участкамі паміж дзвюма негамалагічнымі храмасомамі — транслакацыя (мал. 37.3).

Геномныя мутацыі прыводзяць да змянення колькасці храмасом у клетках. *Асноўнымі прычынамі мутацый такога тыпу з'яўляюцца парушэнні нармальнага працякання меёзу ці мітозу, у выніку якіх разыходжанне храмасом да полюсаў клеткі ажыццяўляецца нераўнамерна або не адбываецца зусім.* Сярод мутацый такога тыпу можна вылучыць гетэраплаідыю, *гаплоідыю * і поліплаідыю (мал. 37.4).

Гетэраплаідыя *(анеуплаідыя)* — гэта змяненне ліку храмасом, не кратнае гаплоіднаму набору. *Такія мутацыі ўзнікаюць, напрыклад, калі ў анафазе мітозу сястрынскія храматыды адной, дзвюх ці некалькіх храмасом не разыходзяцца да полюсаў клеткі ці перамяшчаюцца да аднаго з іх. Падобнае можа адбывацца і пры меёзе: у першым дзяленні — з гамалагічнымі храмасомамі, у другім — з сястрынскімі храматыдамі (мал. 37.5). У выніку ўтвараюцца даччыныя клеткі з недахопам ці, наадварот, лішкам пэўных храмасом.*


Пры гетераплаідыі можа назірацца, напрыклад, адсутнасць у карыятыпе адной з храмасом — монасамія (2n – 1) ці пары гамалагічных храмасом — нулісамія (2n – 2), або наяўнасць у наборы лішніх гамалагічных храмасом — полісамія. Прыкладамі апошняй могуць служыць трысамія (2n + 1), тэтрасамія (2n + 2), пентасамія (2n + 3) і г. д.
*Гаплаідыя — гэта змяншэнне храмасомнага набору да гаплоіднага (1n). Поліплаідыя, наадварот, уяўляе сабой павелічэнне колькасці храмасом у клетках, кратнае гаплоідным набору. *Узнікненне гэтых мутацый, як і ў выпадку гетэраплаідыі, звязана перш за ўсё з парушэннем разыходжання храмасом (храматыд) у меёзе ці мітозе.* У залежнасці ад таго, колькі гаплоідных набораў храмасом змяшчаецца ў клетках, адрозніваюць трыплаідыю (3n), тэтраплаідыю (4n), пентаплаідыю (5n), гексаплаідыю (6n) і г. д. Поліплаідыя распаўсюджана пераважна сярод раслін. Паліплоідныя расліны звычайна маюць больш буйныя вегетатыўныя і генератыўныя органы, чым у дыплоідных форм, адрозніваюцца павышанай устойлівасцю да неспрыяльных фактараў асяроддзя.
● *У залежнасці ад выніку (значэння) для арганізма адрозніваюць наступныя тыпы мутацый. Лятальныя мутацыі прыводзяць да гібелі арганізма. Напрыклад, у чаловека адсутнасць Х-храмасом у наборы выклікае гібель плода на трэцім месяцы эмбрыянальнага развіцця. Паўлятальныя мутацыі зніжаюць жыццяздольнасць мутантаў (гемафілія, прыроджаная форма цукровага дыябету і інш.). Нейтральныя мутацыі не аказваюць істотнага ўплыву на жыццяздольнасць і пладавітасць асобін, напрыклад з'яўленне вяснушак. Карысныя мутацыі павялічваюць прыстасаванасць арганізмаў да ўмоў асяроддзя. Прыкладам могуць служыць мутацыі, якія абумоўліваюць неўспрымальнасць чалавека да пэўных узбуджальнікаў захворванняў — ВІЧ, туберкулёзнай палачкі і інш.*
*Закон гамалагічных радоў спадчыннай зменлівасці. Важны ўклад у развіццё навукі пра спадчынную зменлівасць зрабіў вучоны М. І. Вавілаў. Шмат гадоў ён прысвяціў даследаванню культурных раслін і іх дзікіх продкаў, вывучыў тысячы сартоў. На аснове гэтых даследаванняў М. І. Вавілаў выявіў важную заканамернасць у праяўленні мутацый у роднасных відаў, вядомую як закон гамалагічных радоў спадчыннай зменлівасці. Сутнасць закона выяўляецца ў тым, што віды і роды, блізкія генетычна, звязаныя адзінствам паходжання, характарызуюцца падобнымі радамі спадчыннай зменлівасці. Ведаючы, якія формы зменлівасці сустракаюцца ў аднаго віду, можна прадбачыць знаходжанне падобных форм у роднасных яму відаў.
Хоць даследаванні М. І. Вавілава датычыліся зменлівасці раслін, сфармуляваны ім закон аказаўся ўніверсальным для жывой прыроды. У цяпершні час гамалагічныя рады мутацый выяўлены ў прадстаўнікоў самых розных груп арганізмаў. Прыкладамі могуць служыць выпадкі альбінізму ў розных пазваночных жывёл, прыроджанай формы цукровага дыябету, гемафіліі ці адсутнасці шэрсці ў розных відаў млекакормячых, чырвонай афарбоўкі зярнят ці адсутнасці асцюкоў у суквеццях злакаў і г. д. Узнікненне падобных мутацый у роднасых відаў абумоўлена блізкасцю іх генатыпаў. Пры гэтым, чым большая ступень эвалюцыйнай роднасці арганізмаў, тым большае падабенства назіраецца ў радах спадчыннай зменлівасці.
Закон М. І. Вавілава мае важнае значэнне для селекцыйнай практыкі і сельскай гаспадаркі, бо дазваляе прагназаваць пошук пэўных форм зменлівасці ў раслін, жывёл і мікраарганізмаў. Ведаючы характар спадчыннай зменлівасці таго ці іншага віду, можна прадказваць, якія мутацыі павінны выяўляцца ў роднасных відаў. Дзякуючы закону гамалагічных радоў медыцына і ветэрынарыя атрымалі магчымасць пераносіць веды пра механізмы развіцця, ход і спосабы лячэння спадчынных захворванняў адных відаў (у прыватнасці, чалавека) на іншыя, блізкароднасныя.*
Значэнне генатыпічнай зменлівасці. Спадчынная (генатыпічная) зменлівасць з’яўляецца галоўнай прычынай дзівоснай разнастайнасці жывых арганізмаў на Зямлі. Мутацыйная і камбінатыўная зменлівасць спрыяюць з’яўленню новых спадчынных прымет і іх спалучэнняў і такім чынам абумоўліваюць узнікненне індывідуальных адрозненняў паміж асобінамі. Гэта ляжыць у аснове выжывання і размнажэння адных арганізмаў, больш адаптаваных да ўмоў навакольнага асяроддзя, і гібелі другіх, менш прыстасаваных. Іншымі словамі, спадчынная зменлівасць пастаўляе матэрыял для натуральнага адбору, што забяспечвае эвалюцыю жывой прыроды.
У той жа час мутацыйная і камбінатыўная зменлівасць спрыяюць узнікненню ў арганізмаў такіх прымет і іх камбінацый, якія выкарыстоўваюцца чалавекам пры вывядзенні новых парод жывёл, сартоў раслін і штамаў мікраарганізмаў. Такім чынам, генатыпічная зменлівасць адыгрывае выключна важную ролю ў селекцыйнай рабоце.

Да генатыпічнай (спадчыннай) зменлівасці належаць камбінатыўная і мутацыйная. Камбінатыўная зменлівасць абумоўлена ўзнікненнем у патомства новых спалучэнняў бацькоўскіх генаў. Крыніцамі камбінатыўнай зменлівасці з’яўляюцца кросінговер і незалежнае разыходжанне храмасом у меёзе, выпадковае зліццё гамет пры апладненні. Прычынай мутацыйнай зменлівасці з’яўляюцца наследуемыя змяненні генетычнага матэрыялу арганізмаў — мутацыі. Яны ўзнікаюць раптоўна, не ўтвараюць неперарыўных радоў зменлівасці, не маюць нормы рэакцыі, з’яўляюцца нявызначанымі (непрадказальнымі) і праяўляюцца індывідуальна. Паводле тыпу клетак, якія муціравалі, адрозніваюць генератыўныя і саматычныя мутацыі, паводле прычыны ўзнікнення — спантанныя і індуцыраваныя, паводле ўзроўню змянення генетычнага матэрыялу — генныя, храмасомныя і геномныя. У залежнасці ад выніку (значэння) для арганізма мутацыі бываюць лятальнымі, паўлятальнымі, нейтральнымі і карыснымі. Блізкароднасныя віды характарызуюцца падобнымі формамі спадчыннай зменлівасці, што абумоўлена агульнасцю іх генатыпаў. Спадчынная зменлівасць дастаўляе матэрыял для натуральнага адбору, забяспечваючы эвалюцыю жывой прыроды, і адыгрывае найважнейшую ролю ў селекцыйнай практыцы.
 |
1. Што ўяўляе сабой камбінатыўная зменлівасць? Прывядзіце прыклады. Назавіце крыніцы камбінатыўнай зменлівасці. 2. Дайце азначэнне паняццяў «мутацыя», «мутагенез», «мутаген». На якія групы прынята дзяліць мутагены? Прывядзіце прыклады. 3. Чым саматычныя мутацыі адрозніваюцца ад генератыўных? 4. Ахарактарызуйце асноўныя тыпы генных, храмасомных і геномных мутацый. 5. Якія тыпы мутацый адрозніваюць паводле прычыны іх узнікнення? Паводле значэння для арганізма? 6. У чым заключаецца падабенства камбінатыўнай і мутацыйнай зменлівасці? У чым заключаецца адрозненне паміж гэтымі відамі зменлівасці? Чым мутацыйная зменлівасць адрозніваецца ад мадыфікацыйнай? 7. Сфармулюйце закон гамалагічных радоў спадчыннай зменлівасці. Чым тлумачыцца ўзнікненне падобных мутацый у роднасных відаў? У чым заключаецца практычнае значэнне закона гамалагічных радоў спадчыннай зменлівасці? 8. У пшаніцы-адназярнянкі гаметы змяшчаюць па 7 храмасом. Колькі храмасом змяшчаецца ў саматычных клетках мутантаў пшаніцы-адназярнянкі, калі да ўзнікнення мутантнай формы прывяла нулісамія? Монасамія? Трыплаідыя? Трысамія? Тэтраплаідыя? Тэтрасамія? 9*. Чорная афарбоўка шэрсці ў кошак дамінуе над рыжай, гетэразіготныя кошкі валодаюць чарапахавай афарбоўкай — чорныя плямы чаргуюцца з рыжымі. Гены, якія кантралююць колер шэрсці, размешчаны ў Х-храмасоме. Тэарэтычна катоў, г. зн. самцоў, з чарапахавай афарбоўкай не павінна быць (чаму?), аднак часам яны нараджаюцца. Як растлумачыць гэту з’яву? Як вы лічыце, якія яшчэ асаблівасці, акрамя незвычайнай афарбоўкі, характэрны для чарапахавых катоў? |