*§ 33-1. Узаемадзеянне неалельных генаў
| Сайт: | Профильное обучение |
| Курс: | Біялогія. 11 клас |
| Книга: | *§ 33-1. Узаемадзеянне неалельных генаў |
| Напечатано:: | Гость |
| Дата: | Пятница, 4 Июль 2025, 04:21 |
Прыметы, наследаванне якіх мы разглядалі раней, кантралююцца генамі адной алельнай пары. Аднак многія прыметы жывых арганізмаў вызначаюцца ўзаемадзеяннем двух ці больш за пар генаў, г. зн. неалельнымі генамі. Асноўнымі тыпамі ўзаемадзеяння неалельных генаў з'яўляюцца камплементарнасць, эпістаз і полімерыя.
Камплементарнасць выяўляецца ў тым, што дамінантныя гены розных алеляў супольна абумоўліваюць з'яўленне новай прыметы, якая не вызначалася ніводным з гэтых генаў паасобку. Напрыклад, у льну дамінантны ген адной алелі (А) вызначае ружовую афарбоўку кветак, а дамінантны ген другой алелі (В) — белую. Калі ў генатыпе расліны прысутнічаюць абодва дамінантныя алелі, яго кветкі маюць блакітную афарбоўку. Такім чынам, неалельныя гены А і В дзейнічаюць камплементарна (узаемна дапаўняюць адзін аднаго), што прыводзіць да фарміравання новай прыметы. Сутнасць такога ўзаемадзеяння можна адлюстраваць у выглядзе схемы.

Як бачна са схемы, сінтэз блакітнага пігменту з неафарбаванага папярэдніка (прапігменту) ажыццяўляецца ў два этапы, кожны з якіх каталізуецца асаблівым ферментам. Наяўнасць гэтых ферментаў у клетках пялёсткаў лёну вызначаецца наяўнасцю ў генатыпе адпаведных генаў А і В. Так, ген А вызначае наяўнасць ферменту, які ператварае прапігмент у прамежкавы прадукт — пігмент ружовага колеру. Ген В абумоўлівае наяўнасць другога ферменту, які сінтэзуе блакітны пігмент з ружовага. Таму ў раслін, што маюць абодва дамінантныя алелі, у пялёстках утвараецца канчатковы прадукт — пігмент блакітнага колеру. Калі адсутнічае ген В (а значыць, і адпаведны фермент), у клетках працякае толькі першая стадыя — утварэнне ружовага пігменту. У выпадку адсутнасці гена А (ці абодвух дамінантных алеляў) не ажыццяўляецца нават першы этап сінтэзу пігменту, і пялёсткі застаюцца белымі.
Разгледзім скрыжаванне дыгомазіготных раслін лёну, якія маюць ружовыя (AAbb) і белыя (aaBB) кветкі (мал. 33-1.1). У першым пакаленні назіраецца аднастайнасць гібрыдаў. Усе яны маюць блакітныя кветкі, што абумоўлена наяўнасцю ў генатыпе як гена А, так і гена В. У другім гібрыдным пакаленні назіраецца расшчапленне ў суадносінах 9 блакітных : 3 ружовых : 4 белых.

Камплементарнае ўзаемадзеянне генаў ляжыць у аснове праяўлення шэрага прымет у розных арганізмаў. Так, па тыпе камплементарнасці ўзаемадзейнічаюць гены, што кантралююць афарбоўку коканаў у тутавага шаўкапрада і кветак у духмянага гарошку, форму грэбеня ў курэй, колер апярэння ў папугаяў і г. д. У залежнасці ад асаблівасцей фенатыпічнага праяўлення генаў у патомства ад скрыжавання дыгетэразігот могуць назірацца розныя варыянты расшчаплення — 9 : 7, 9 : 3 : 4, 9 : 6 : 1, 9 : 3 : 3 : 1.
Напрыклад, у гарбуза дамінантныя алелі А і В супольна прыводзяць да з'яўлення пладоў дыскападобнай формы, а паасобку — шарападобнай. У той жа час рэцэсіўныя алелі гэтых генаў абумоўліваюць падоўжаную форму пладоў. Пры скрыжаванні дыгетэразіготных раслін, якія маюць дыскападобныя плады, у патомстве адбываецца расшчапленне 9 : 6 : 1.
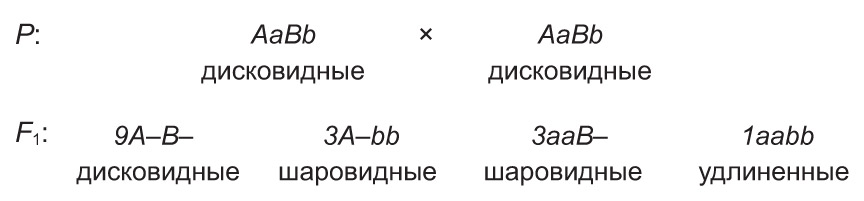
Падобным чынам наследуецца колер апярэння хвалістых папугаяў. Дамінантны ген адной алелі (А) абумоўлівае ў іх блакітную афарбоўку пёраў, а дамінантны ген другой алелі (В) — жоўтую. Пры наяўнасці ў генатыпе абодвух дамінантных алеляў (А і В) выяўляецца новая прымета — зялёнае апярэнне. Разам з тым рэцэсіўныя алелі гэтых генаў (a і b) вызначаюць белую афарбоўку пёраў. Таму скрыжаванне зялёных дыгетэразіготных асобін прыводзіць да расшчаплення па колеры апярэння сярод нашчадкаў у суадносінах 9 : 3 : 3 : 1.
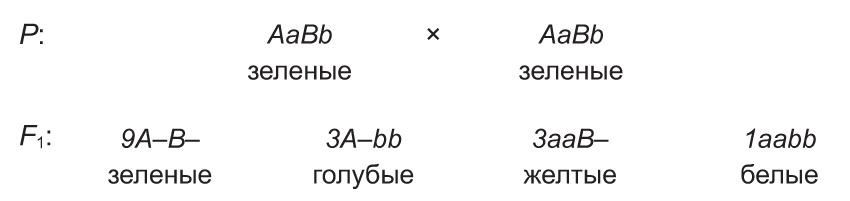
Эпістаз — тып узаемадзеяння, пры якім ген адной алелі прыгнятае праяўленне генаў другой алельнай пары. Такія гены, якія блакіруюць фенатыпічнае праяўленне іншых неалельных генаў, называюцца эпістатычнымі, генамі-інгібітарамі ці супрэсарамі. Гены, якія прыгнятаюцца, у сваю чаргу, атрымалі назву гіпастатычных. У залежнасці ад таго, дамінантным ці рэцэсіўным з'яўляецца ген-інгібітар, адрозніваюць дамінантны і рэцэсіўны эпістаз.
Прыкладам дамінантнага эпістазу можа служыць узаемадзеянне двух алеляў, якія вызначаюць масць (афарбоўку) коней. Так, дамінантны ген адной алельнай пары (А) абумоўлівае варанýю (чорную) масць каня, а рэцэсіўны ген (а) — рыжую. Аднак гэтыя гены могуць выявіцца фенатыпічна толькі пры адсутнасці ў генатыпе каня дамінантнага гена іншай алелі — інгібітару (I). Пры наяўнасці інгібітару праяўленне генаў вараной і рыжай масці прыгнятаецца. У разглядаемым прыкладзе ген-супрэсар мае ўласнае фенатыпічнае праяўленне: у выпадку яго прысутнасці ў асобін развіваецца шэрая афарбоўка.
Разгледзім скрыжаванне дыгетэразіготных коней (мал. 33-1.2). Яны маюць шэрую масць, што абумоўлена наяўнасцю ў генатыпе дамінантнага гена-супрэсара (I). У патомства ад скрыжавання назіраецца расшчапленне 12 : 3 : 1. Усе асобіны, якія змяшчаюць у генатыпе эпістатычны ген (9A—I— і 3aaI—), наследуюць шэрую масць. У жарабят, якія не маюць гена I, выяўляецца вараная або рыжая афарбоўка ў суадносінах 3 вараныя (3A–ii) да 1 рыжай (1aaii).

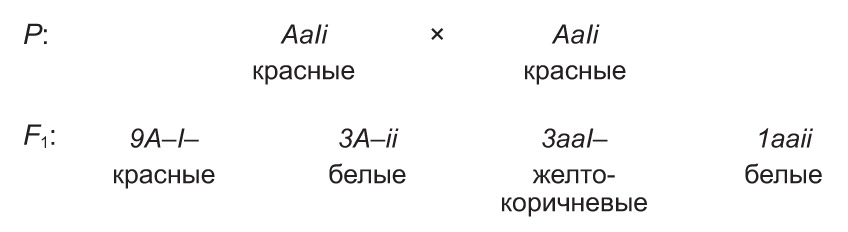
Калі ген-супрэсар з'яўляецца рэцэсіўным, то ўзаемадзеянне неалельных генаў працякае па тыпе рэцэсіўнага эпістазу (крыптамерыі). Такое ўзаемадзеянне назіраецца, напрыклад, пры наследаванні колеру насення фасолі. У гэтай расліны чырвоная афарбоўка насення (А) дамінуе над жоўта-карычневай (а). Праяўленне рэцэсіўнага інгібітару (i) вядзе да блакіравання дзеяння генаў чырвонай і жоўта-карычневай афарбоўкі, і насенне аказваецца белым. Аднак дзеянне рэцэсіўнага эпістатычнага гена можа выявіцца толькі пры адсутнасці ў генатыпе адпаведнага яму дамінантнага алеля (I). У выпадку наяўнасці такога гена дзеянне супрэсара прыглушаецца. Так, скрыжаванне дыгетэразіготных раслін фасолі прыводзіць да расшчаплення сярод нашчадкаў у суадносінах:  з чырвоным насеннем,
з чырвоным насеннем,  з жоўта-карычневымі і
з жоўта-карычневымі і  з белымі.
з белымі.
Эпістаз назіраецца пры наследаванні многіх прымет жывых арганізмаў. Пры розных формах эпістазу і розных асаблівасцях праяўлення генаў расшчапленне ў патомстве ад скрыжавання дыгетэразіготных асобін можа складаць 13 : 3, 12 : 3 : 1 або 9 : 3 : 4.
У чалавека апісаны так званы «бамбейскі феномен». Сутнасць яго заключаецца ў тым, што ў некаторых людзей, якія маюць у генатыпе ген IA ці IB, фенатыпічна выяўляецца першая (0) група крыві. Было вызначана, што ў аснове гэтай з'явы ляжыць рэцэсіўны эпістаз. Дадзеныя людзі аказаліся гомазіготнымі па рэдкім рэцэсіўным гене, што парушае сінтэз антыгенаў А і В. Таму незалежна ад спалучэння генаў, якія вызначаюць групу крыві па сістэме АВ0, эрытрацыты такіх людзей не змяшчаюць антыгенаў А і В.
Полімерыя — узаемадзеянне, пры якім гены двух ці больш алеляў праяўляюцца падобным чынам, вызначаючы развіццё адной і той жа прыметы. У выпадку полімерыі неалельныя гены, якія кантралююць тую ці іншую прымету (палімерныя гены), прынята абазначаць аднолькавымі літарамі з лічбавымі індэксамі. Пры гэтым гены кожнай алельнай пары маюць аднолькавы індэкс. Напрыклад: А1, а1 — дамінантны і рэцэсіўны алелі першай пары генаў, А2, а2 — другой пары і г. д. Вядомыя дзве разнавіднасці полімерыі.
Пры некумулятыўнай полімерыі дамінантная прымета выяўляецца ў поўнай меры пры наяўнасці ў генатыпе хаця б аднаго дамінантнага гена. Іншымі словамі, колькасць дамінантных генаў не ўплывае на ступень выяўленасці прыметы. Так, у стрэлак форма пладоў вызначаецца дзвюма парамі палімерных генаў. Пры гэтым рэцэсіўная прымета — авальныя плады — выяўляецца толькі ў раслін, якія маюць генатып а1а1а2а2. Калі ў генатыпе прысутнічае хаця б адзін дамінантны ген (няважна, які менавіта — А1 ці А2), расліна валодае дамінантнай прыметай — пладамі трохвугольнай формы.
Скрыжаванне дамінантнай і рэцэсіўнай дыгомазігот (мал. 33-1.3) прыводзіць да з'яўлення ў першым пакаленні дыгетэразіготных гібрыдаў з трохвугольнымі пладамі (А1а1А2а2). У другім пакаленні назіраецца расшчапленне ў суадносінах 15 : 1.

У выпадку кумулятыўнай полімерыі ступень выяўленасці дамінантнай прыметы вызначаецца колькасцю дамінантных генаў у генатыпе. Чым больш такіх генаў, тым мацней выяўляецца дадзеная прымета. Напрыклад, у пшаніцы пэўных сартоў афарбоўка зярнят кантралюецца трыма парамі неалельных генаў. Расліны з генатыпам а1а1а2а2а3а3 маюць белыя зярняты, а тыя, што валодаюць генатыпам А1А1А2А2А3А3, — цёмна-чырвоныя. У раслін з іншымі генатыпамі назіраюцца розныя прамежкавыя варыянты афарбоўкі, што залежаць ад ліку дамінантных генаў. Так, наяўнасць у генатыпе толькі аднаго дамінантнага гена абумоўлівае бледна-ружовы колер зярнят, двух — ружовы, трох — светла-чырвоны і г. д.
Вызначана, што па тыпе кумулятыўнай полімерыі наследуюцца многія колькасныя прыметы арганізмаў: рост чалавека, колер яго скуры (г. зн. колькасць меланіну), яйцаноскасць курэй, малочнасць кароў, утрыманне цукрозы ў караняплодах цукровых буракоў, бялкоў у эндасперме насення траў і інш.
Некаторыя гены, якія не маюць уласнага фенатыпічнага праяўлення, здольныя ўзмацняць ці, наадварот, аслабляць (у адрозненне ад эпістазу не прыгнятаючы цалкам) праяўленне іншых неалельных генаў. Такія гены называюць мадыфікатарамі, а іх дзеянне — мадыфікуючым. Напрыклад, у кошак чорны колер шэрсці дамінуе над белым. Гетэразіготныя асобіны валодаюць чорнай афарбоўкай, але, як правіла, адзін ці некалькі участкаў іх шэрсці мае белы колер. Колькасць і памер такіх участкаў залежаць ад адпаведных генаў-мадыфікатараў. У шэрых мышэй падобным чынам вызначаецца памер белай плямы на брушку. Мяркуецца, што ў чалавека колер радужнай абалонкі вачэй кантралюецца як мінімум шасцю парамі генаў. Пры гэтым разам з «асноўнымі» генамі, якія вызначаюць карыя і блакітныя вочы, істотны ўплыў на афарбоўку радужкі аказваюць гены-мадыфікатары. Іх дзеянне прыводзіць да з'яўлення разнастайных адценняў — ад светла-шэрага ці светла-блакітнага да практычна чорнага.

Многія прыметы жывых арганізмаў вызначаюцца ўзаемадзеяннем неалельных генаў. Асноўнымі тыпамі такога ўзаемадзеяння з'яўляюцца камплементарнасць, эпістаз і полімерыя. Пры камплементарнам дзеянні дамінантныя гены розных алеляў разам абумоўліваюць з'яўленне новай прыметы, якая не вызначалася ні адным з гэтых генаў паасобку. У выпадку эпістаза ген адной алелі (эпістатычны ген) падаўляе праяўленне генаў другой алельнай пары. У залежнасці ад таго, дамінантным або рэцэсіўным з'яўляецца эпістатычны ген, адрозніваюць дамінантны і рэцэсіўны эпістаз. Полімерыя заключаецца ў тым, што гены двух або больш алеляў выяўляюцца падобна, вызначаючы развіццё адной і той жа прыметы. Пры кумулятыўнай полімерыі ступень праявы дамінантнай прыметы залежыць ад колькасці дамінантных генаў, пры некумулятыўнай наяўнасць нават аднаго дамінантнага гена вядзе да поўнай праявы дамінантнай прыметы.
 |
1. Пакажыце тыпы ўзаемадзеяння неалельных генаў: кадамінаванне, эпістаз, полімерыя, поўнае дамінаванне, няпоўнае дамінаванне, камплементарнасць. 2. Што ўяўляе сабой камплементарнае ўзаемадзеянне неалельных генаў? Прывядзіце прыклады. 3. У чым заключаецца дзеянне эпістатычных генаў? Чым дамінантны эпістаз адрозніваецца ад рэцэсіўнага? 4. Што такое полімерыя? У чым выражаецца адрозненне паміж кумулятыўнай і некумулятыўнай полімерыяй? 5. Якія варыянты расшчаплення могуць назірацца ў патомстве ад скрыжавання дыгетэрозігот ў выпадку ўзаемадзеяння неалельных генаў па тыпу: а) камплементарнасці; б) эпістаза; в) некумулятыўнай полімерыі? 6*. У хамякоў карычневая афарбоўка шэрсці вызначаецца адным дамінантным генам, а абрыкосавая — другім. Дадзеныя гены размешчаны ў розных парах храмасом, іх рэцэсіўныя алелі прыводзяць да з'яўлення белай афарбоўкі. Калі ж у генатыпе хамяка прысутнічаюць абодва неаллельных дамінантных гена, з'яўляецца новая прымета — чорная афарбоўка. Ад скрыжавання абрыкосавага самца з карычневай самкай нарадзіўся хамяк з белай поўсцю. Устанавіце генатыпы бацькоўскіх асобін. Якое патомства можна чакаць у іх у далейшым? 7*. Афарбоўка шэрсці сабак пароды лабрадор кантралюецца двума алелямі, лакалізаванымі ў розных парах храмасом. Гены адной алельнай пары абумоўліваюць развіццё чорнай або карычневай афарбоўкі, пры гэтым чорная масць цалкам дамінуе над карычневай. У другой алельнай пары рэцесіўны ген валодае эпістатычным дзеяннем. Ён падаўляе праяўленне генаў чорнай і карычневай афарбоўкі, абумоўлівая развіццё шэрсці светла-бэжавага колеру. Дыгетэрозіготную асобіну скрыжавалі з рэцэсіўнай дыгомазіготнай. Які фенатып маюць скрыжаваныя асобіны? Якая верагоднасць з'яўлення ў іх патомства шчанюкоў, якія маюць: а) чорную; б) карычневую; в) светла-бэжавую шэрсць? 8*. Дапусцім, што ў раслін пэўнага віду даўжыня пялёсткаў вызначаецца двума парамі палімерных генаў, размешчаных ў негамалагічных храмасомах. Асобіны з даўжынёй пялёсткаў 40 мм маюць цалкам дамінантны генатып, а з даўжынёй 20 мм — цалкам рэцэсіўны. Расліны з іншымі генатыпамі маюць пялёсткі даўжынёй 35, 30 або 25 мм. Якая даўжыня пялёсткаў у дыгетэразіготных раслін? Якое расшчапленне ў патомства будзе назірацца пры скрыжаванні гэтых раслін паміж сабой? |