§ 31. Цыталагічныя асновы наследавання прымет пры монагібрыдным скрыжаванні
Паняцце пра алельныя гены. Цыталагічныя асновы наследавання прымет пры монагібрыдным скрыжаванні. Толькі пасля таго, як былі адкрыты храмасомы, апісаны іх паводзіны пры мітозе і меёзе і даказана, што гены размяшчаюцца ў храмасомах, меркаванні Г. Мендэля знайшлі навуковае пацверджанне.
Гены, якія кантралююць розныя (альтэрнатыўныя) формы праяўлення прыметы, называюцца алельными генамі ці алелямі. Вызначана, што алельныя гены размяшчаюцца ў аднолькавых участках (локусах) гамалагічных храмасом. Такім чынам, у любога дыплоіднага арганізма праяўленне той ці іншай прыметы вызначаецца як мінімум двума алельнымі генамі.
Сукупнасць усіх генаў арганізма называюць генатыпам. У дачыненні да асобнай прыметы словам «генатып» абазначаюць камбінацыю алельных генаў, якія кантралююць дадзеную прымету. Арганізмы, якія маюць аднолькавыя алельныя гены, называюцца гомазіготамі. Адрозніваюць дамінантныя гомазіготы (іх генатып можна запісаць як АА) і рэцэсіўныя гомазіготы (аа). Асобіны, якія маюць розныя алельныя гены, называюцца гетэразіготамі, іх генатып можна абазначыць як Аа.
Гаметы ўтвараюцца ў выніку меёзу і змяшчаюць гаплоідны набор храмасом. Успомнім, што ў анафазе I гамалагічныя храмасомы, якія змяшчаюць алельныя гены, разыходзяцца да процілеглых полюсаў клеткі, што дзеліцца, і ў канчатковым выніку трапляюць у розныя гаметы (мал. 31.2). Такім чынам, два алельныя гены не могуць апынуцца ў адной і той жа палавой клетцы. У кожную гамету трапляе толькі адзін з іх.

*Гіпотэзу пра тое, што алельныя гены размяркоўваюцца пароўну паміж палавымі клеткамі, не трапляючы ў адну гамету, не разбаўляючыся і не змешваючыся адзін з адным, упершыню выказаў англійскі генетык У. Бэтсан у 1902 г. Сваё меркаванне вучоны назваў гіпотэзай чысціні гамет. Пасля гэтага гіпотэза была цалкам пацверджана данымі цыталогіі і генетыкі. Таму ў цяперашні час прынцып, сфармуляваны У. Бэтсанам, называюць не гіпотэзай, а правілам чысціні гамет.*
Гомазіготныя арганізмы маюць аднолькавыя алельныя гены, таму ў іх фарміруецца толькі адзін тып гамет. У асобін з генатыпам АА ўсе палавыя клеткі змяшчаюць ген А, у арганізмаў з генатыпам аа ўсе гаметы змяшчаюць ген а. Гетэразіготныя асобіны (Аа) утвараюць два тыпы гамет у роўных суадносінах: 50 % палавых клетак змяшчаюць алель А, 50 % — алель а.
Пры апладненні гаплоідныя гаметы бацькоў зліваюцца з утварэннем дыплоіднай зіготы. У зігоце храмасомы зноў становяцца парнымі. У кожнай пары гамалагічных храмасом адна з’яўляецца мацярынскай, а другая — бацькоўскай. Значыць, у кожнага нашчадка развіццё той ці іншай прыметы будзе вызначацца двума алельными генамі, прычым адзін з іх унаследаваны ад маці, а другі — ад бацькі.
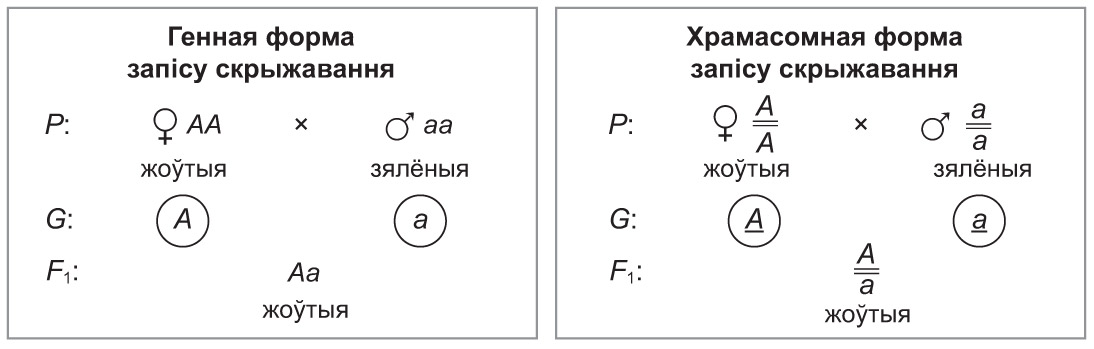
Вернемся да эксперымента, у якім Г. Мендэль вывучаў наследаванне афарбоўкі насення гароху. Абазначым дамінантны ген, які абумоўлівае жоўтую афарбоўку, літарай А, і рэцэсіўны ген, што вызначае зялёную афарбоўку, — а. Паколькі Г. Мендэль выкарыстоўваў у якасці бацькоўскіх форм асобін чыстых ліній, іх генатыпы трэба запісаць як АА і аа. Абодва бацькі — гомазіготы, кожны з іх утварае гаметы толькі аднаго тыпу: у асобіны з генатыпам АА фарміруюцца толькі гаметы А, у асобіны з генатыпам аа — толькі гаметы а.
Зліццё гамет прывяло да ўтварэння зігот, з якіх развіліся гібрыды першага пакалення. Відавочна, што ўсе яны мелі генатып Аа і жоўтую афарбоўку насення (дамінантны ген цалкам падавіў праяўленне рэцэсіўнага).
Запішам дадзенае монагібрыднае скрыжаванне. Найбольш распаўсюджанымі формамі запісу скрыжаванняў з’яўляюцца генная і храмасомная. У першым выпадку гены запісваюць «у радок», без указання храмасом (напрыклад, Аа). У другім выпадку пры запісе генатыпаў алельныя гены размяшчаюць адзін над адным. Пры гэтым дзвюма рыскамі абазначаюць гамалагічныя храмасомы, у якіх гэтыя гены змяшчаюцца (напрыклад,  ). Тут і далей выкарыстоўвайце адну з форм запісу (па ўказанні настаўніка).
). Тут і далей выкарыстоўвайце адну з форм запісу (па ўказанні настаўніка).
Сукупнасць прымет і ўласцівасцей арганізма называюць фенатыпам. Калі гаворка ідзе пра пэўнае скрыжаванне, паняццем «фенатып» абазначаюць тую прымету (ці прыметы), якая ў гэтым скрыжаванні даследуецца. Напрыклад, у разгледжаным выпадку можна сказаць, што гібрыды першага пакалення мелі аднолькавы фенатып — жоўты колер насення.
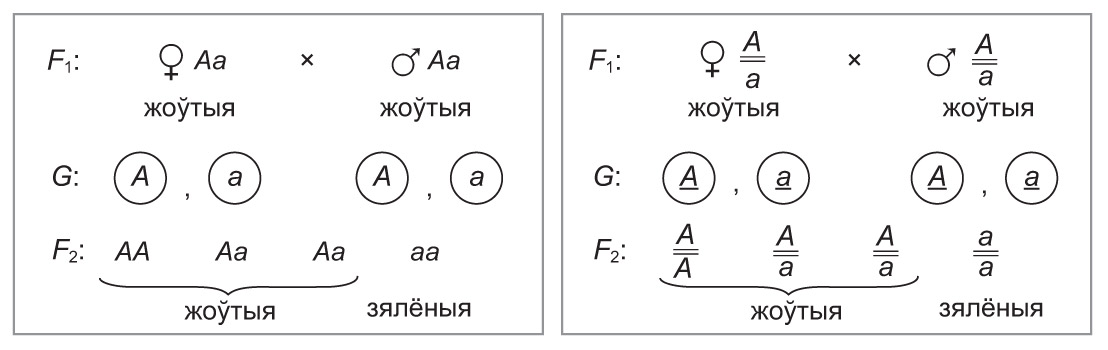
 Гібрыды першага пакалення — гетэразіготы (Аа), у іх утвараюцца два тыпы гамет (А і а) у роўных суадносінах. Зліццё гамет носіць выпадковы характар, таму пры апладненні фарміруюцца розныя тыпы зігот: АА, Аа і аа.
Гібрыды першага пакалення — гетэразіготы (Аа), у іх утвараюцца два тыпы гамет (А і а) у роўных суадносінах. Зліццё гамет носіць выпадковы характар, таму пры апладненні фарміруюцца розныя тыпы зігот: АА, Аа і аа.
Каб наглядна паказаць усе варыянты зліцця гамет і разлічыць верагоднасць з’яўлення патомства з рознымі генатыпамі і фенатыпамі, можна пабудаваць табліцу, якая называецца рашоткай Пенэта (упершыню такія табліцы прапанаваў выкарыстоўваць англійскі генетык Р. Пенэт). У рашотцы Пенэта па гарызанталі запісваюць гаметы аднаго з бацькоў, па вертыкалі — гаметы другога. У клетках на скрыжаванні радкоў і слупкоў паказваюць генатыпы і фенатыпы асобін, якія ўзнікаюць пры зліцці адпаведных гамет (мал. 31.3).
Як бачна з пабудаванай рашоткі, у гетэразіготных бацькоўскіх форм утвараецца патомства з трыма генатыпамі ў суадносінах: 1АА : 2Аа : 1аа. Такім чынам, расшчапленне па генатыпе складае 1 : 2 : 1. Верагоднасць з’яўлення нашчадкаў кожнага тыпу можна паказаць і ў працэнтах : 25 % АА, 50 % Аа, 25 % аа.
Расшчапленне па прымеце, якая даследуецца, такое:  асобін з жоўтым насеннем (75 %) і
асобін з жоўтым насеннем (75 %) і  — з зялёным (25 %). Значыць, расшчапленне па фенатыпе складае 3 : 1.
— з зялёным (25 %). Значыць, расшчапленне па фенатыпе складае 3 : 1.
Хоць расліны з жоўтым насеннем знешне выглядаюць аднолькава, генетычна яны неаднастайныя (АА і Аа). Становяцца зразумелымі прычыны розных «паводзін» іх патомства ў трэцім і наступных пакаленнях. Пры самаапыленні сярод нашчадкаў дамінантных гомазігот АА не будзе назірацца расшчаплення, як і сярод нашчадкаў рэцэсіўных гомазігот аа. Гетэразіготныя асобіны Аа будуць даваць у патомстве расшчапленне 3 : 1.
Такім чынам, цыталагічнымі асновамі першага і другога законаў Мендэля з’яўляюцца разыходжанне гамалагічных храмасом у анафазе I меёзу і выпадковае зліццё (спалучэнне) гамет пры апладненні.